6.2 Storage: Maintaining Memories over Time
Encoding is the process of turning perceptions into memories. But one of the hallmarks of a memory is that you can bring it to mind on Tuesday, not on Wednesday, and then bring it to mind again on Thursday. So where are our memories when we are not using them? Clearly, those memories are stored in some form in your brain. As pointed out earlier, storage is the process of maintaining information in memory over time. There are three major kinds of memory storage: sensory, short-term, and long-term. As these names suggest, the three kinds of storage are distinguished primarily by the amount of time over which a memory is retained.
6.2.1 Sensory Storage

 Figure 6.5: Iconic Memory Test When a grid of letters is flashed on screen for only 1/20th of a second, it is difficult to recall individual letters. But if prompted to remember a particular row immediately after the grid is shown, research participants will do so with high accuracy. Sperling used this procedure to demonstrate that although iconic memory stores the whole grid, the information fades away too quickly for a person to recall everything (Sperling, 1960).
Figure 6.5: Iconic Memory Test When a grid of letters is flashed on screen for only 1/20th of a second, it is difficult to recall individual letters. But if prompted to remember a particular row immediately after the grid is shown, research participants will do so with high accuracy. Sperling used this procedure to demonstrate that although iconic memory stores the whole grid, the information fades away too quickly for a person to recall everything (Sperling, 1960).
Sensory memory is a type of storage that holds sensory information for a few seconds or less. In a series of classic experiments, research participants were asked to remember rows of letters (Sperling, 1960). In one version of the procedure, participants viewed three rows of four letters each, as shown in FIGURE 6.5. The researcher flashed the letters on a screen for just 1/20th of a second. When asked to remember all 12 of the letters they had just seen, participants recalled fewer than half. There were two possible explanations for this: Either people simply could not encode all the letters in such a brief period of time, or they had encoded the letters but forgotten them while trying to recall everything they had seen.
To test the two ideas, the researcher relied on a clever trick. Just after the letters disappeared from the screen, a tone sounded that cued the participants to report the letters in a particular row. A high tone cued participants to report the contents of the top row, a medium tone cued participants to report the contents of the middle row, and a low tone cued participants to report the contents of the bottom row. When asked to report only a single row, people recalled almost all of the letters in that row! Because the tone sounded after the letters disappeared from the screen, the researchers concluded that people could have recalled the same number of letters from any of the rows, had they been asked. Participants had no way of knowing which of the three rows would be cued, so the researcher inferred that virtually all the letters had been encoded. In fact, if the tone was substantially delayed, participants could not perform the task because the information had slipped away from their sensory memories. Like the afterimage of a flashlight, the 12 letters flashed on a screen are visual icons, a lingering trace stored in memory for a very short period.
How long is information held in iconic and echoic memory before it decays?
Because we have more than one sense, we have more than one kind of sensory memory. Iconic memory is a fast-decaying store of visual information. A similar storage area serves as a temporary warehouse for sounds. Echoic memory is a fast-decaying store of auditory information. When you have difficulty understanding what someone has just said, you probably find yourself replaying the last few words—listening to them echo in your “mind’s ear,” so to speak. When you do that, you are accessing information that is being held in your echoic memory store. The hallmark of both the iconic and echoic memory stores is that they hold information for a very short time. Iconic memories usually decay in about 1 second or less, and echoic memories usually decay in about 5 seconds (Darwin, Turvey, & Crowder, 1972). These two sensory memory stores are a bit like doughnut shops: The products come in, they sit briefly on the shelf, and then they are discarded. If you want one, you have to grab it fast.
Page 229
6.2.2 Short-Term Storage and Working Memory
A second kind of memory storage is short-term memory, which holds nonsensory information for more than a few seconds but less than a minute. For example, if someone tells you a telephone number, you can usually repeat it back with ease—but only for a few seconds. In one study, research participants were given consonant strings to remember, such as DBX and HLM. After seeing each string, participants were asked to count backward from 100 by 3 for varying amounts of time and were then asked to recall the strings (Peterson & Peterson, 1959). As shown in FIGURE 6.6, memory for the consonant strings declined rapidly, from approximately 80 percent after a 3-second delay to less than 20 percent after a 20-second delay. These results suggest that information can be held in the short-term memory store for about 15 to 20 seconds.
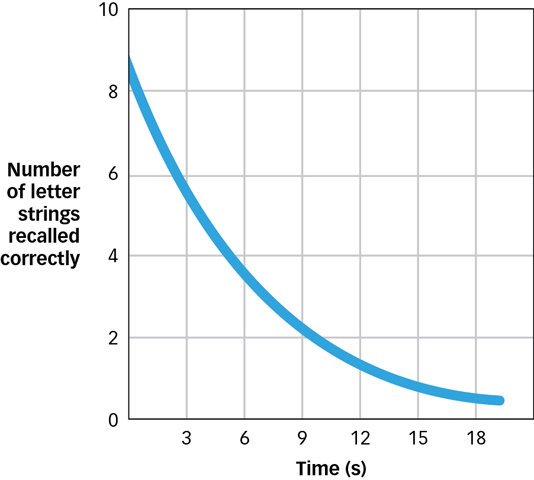
Figure 6.6: The Decline of Short-Term Memory A 1959 experiment showed how quickly short-term memory fades without rehearsal. On a test for memory of three-letter strings, research participants were highly accurate when tested a few seconds after exposure to each string, but if the test was delayed another 15 seconds, people barely recalled the strings at all (Peterson & Peterson, 1959).
Why is it helpful to repeat a telephone number you are trying to remember?
What if 15 to 20 seconds is not enough time? What if we need the information for a while longer? We can use a trick that allows us to get around the natural limitations of our short-term memories. Rehearsal is the process of keeping information in short-term memory by mentally repeating it. If someone gives you a telephone number and you cannot put it immediately into your cell phone or write it down, you say it over and over to yourself until you can. Each time you repeat the number, you are re-entering it into short-term memory, giving it another 15 to 20 seconds of shelf life.
Short-term memory is limited in how long it can hold information, and also limited in how much information it can hold. Not only can most people keep approximately seven numbers in short-term memory, but if they put more new numbers in, then old numbers begin to fall out (Miller, 1956). Short-term memory is not limited to numbers, of course. It can also hold about seven letters or seven words—even though those seven words contain many more than seven letters. In fact, short-term memory can hold about seven meaningful items at once (Miller, 1956). Therefore, one way to increase storage is to group several letters into a single meaningful item. Chunking involves combining small pieces of information into larger clusters or chunks that are more easily held in short-term memory. Servers who use organizational encoding (p. 225) to organize customer orders into groups are essentially chunking the information, giving themselves less to remember.
Short-term memory was originally conceived of as a kind of “place” where information is kept for a limited amount of time. More recently, British researchers developed and refined a more dynamic model of a limited-capacity memory system, working memory, which refers to active maintenance of information in short-term storage (Baddeley & Hitch, 1974). Working memory includes subsystems that store and manipulate visual images or verbal information, as well as a central executive that coordinates the subsystems (Baddeley, 2001). If you wanted to keep the arrangement of pieces on a chessboard in mind as you contemplated your next move, you would be relying on working memory. Working memory includes the visual representation of the positions of the pieces, your mental manipulation of the possible moves, and your awareness of the flow of information into and out of memory, all stored for a limited amount of time. In short, the working memory model acknowledges both the limited nature of this kind of memory storage and the activities that are commonly associated with it.
Research conducted in the context of this model has taught us that working memory plays an important role in many aspects of our cognitive lives. For example, studies of individuals with neurological damage to the verbal subsystem of working memory reveal that, not only do they have problems holding onto strings of digits and letters for a few seconds, they also have difficulty learning novel words. This suggests a link between this part of the working memory system and the ability to learn language (Baddeley, 2001; Gathercole, 2008).
Page 230
Brain imaging studies indicate that the central executive component of working memory depends on regions within the frontal lobe that are important for controlling and manipulating information on a wide range of cognitive tasks (Baddeley, 2001). Children who score low on working memory tasks have difficulty learning new information and performing well in the classroom (Alloway et al., 2009). Can working memory skills be trained, and can such training enhance cognitive functioning? This question has become a hot research topic over the past few years (Klingberg, 2010; Shipstead, Redick, & Engle, 2012).
In typical studies, participants are first given extensive practice performing working memory tasks that require the maintenance and manipulation of visual or verbal information. They are then tested on new working memory tasks that have not been specifically taught, as well as on other cognitive tasks that tap into capacities such as reasoning, comprehension, or sustained attention. Some encouraging results have been reported. For example, elementary school students trained on several working memory tasks (about 35 minutes/day for at least 20 days over a 5- to 7-week time period) showed improvement on other working memory tasks when compared with untrained low working memory children (Holmes, Gathercole, & Dunning, 2009). These gains were evident even when the children were tested 6 months after training. There was also some evidence of improvement on math tasks. However, working memory training studies typically compare a working memory training group with control groups that do not perform any kind of training, or perform less challenging training tasks than the working memory group performs, so it is not clear whether working memory training in particular is responsible for the effects that are observed (Slagter, 2012). Indeed, a recent study that used a control condition involving active processing (a visual search task) found that working memory training improved performance on the working memory task that was trained, but did not result in improvements on other cognitive tasks (Redick et al., 2013). More research will be needed to determine whether working memory training produces any general improvements in cognitive performance (Shipstead et al., 2012).
6.2.3 Long-Term Storage

Even years after leaving home in Pontito, Italy, painter Franco Magnani was able to create a near-perfect reproduction of what he had seen there. Magnani’s painting (left), based on a memory of a place he had not seen for years, is remarkably similar to the photograph (right) Susan Schwartzenberg took of the actual scene.
© FRANCO MAGNANI and SUSAN SCHWARTZENBERG. © EXPLORATORIUM, WWW.EXPLORATORIUM.EDU
The artist Franco Magnani was born in Pontito, Italy, in 1934. In 1958, he left his village to see the rest of the world and settled in San Francisco, California in the 1960s. Soon after arriving, Magnani began to suffer from a strange illness. Every night he experienced feverish dreams of Pontito, in which he recalled the village in vivid detail. The dreams soon penetrated his waking life in the form of overpowering recollections, and Magnani decided that the only way to rid himself of these images was to capture them on canvas. For the next 20 years, he devoted much of his time to painting, in exquisite detail, his memories of his beloved village. Many years later, photographer Susan Schwartzenberg went to Pontito, armed with a collection of Magnani’s paintings, and photographed each scene from the perspective of the paintings. As you can see in the images, the correspondence between the paintings and the photographs was striking (Sacks, 1995; Schacter, 1996).
Page 231
Many years intervened between Magnani’s visual perception and artistic reconstruction of the village, suggesting that very detailed information can sometimes be stored for a very long time. In contrast to the time-limited sensory memory and short-term memory stores, long-term memory is a type of storage that holds information for hours, days, weeks, or years. In contrast to both sensory and short-term memory, long-term memory has no known capacity limits (see FIGURE 6.7). For example, most people can recall 10 000 to 15 000 words in their native language, tens of thousands of facts (The capital of France is Paris and 3 × 3 = 9), and an untold number of personal experiences. Just think of all the song lyrics you can recite by heart, and you will understand that you have a lot of information tucked away in long-term memory!
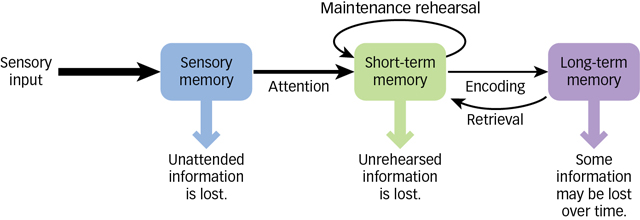
Figure 6.7: The Flow of Information through the Memory System Information moves through several stages of memory as it gets encoded, stored, and made available for later retrieval.
Amazingly, people can recall items from long-term memory even if they have not thought of them for years. For example, researchers have found that even 50 years after graduation, people can accurately recognize about 90 percent of their high school classmates from yearbook photographs (Bahrick, 2000). The feat is more remarkable when you consider that probably most of this information had not been accessed for years before the experiment.
6.2.3.1 The Role of the Hippocampus as Index
Where is long-term memory located in the brain? The clues to answering this question come from individuals who are unable to store long-term memories. Not everyone has the same ability to encode information into long-term memory. In 1953, 27-year-old Henry Molaison suffered from severe epilepsy and was not responding to medication (Scoville & Milner, 1957). In a desperate attempt to stop the seizures, Henry’s surgeon removed parts of his temporal lobes, including the hippocampus and some surrounding regions (FIGURE 6.8). After the operation, Henry could converse easily, use and understand language, and perform well on intelligence tests, but he could not remember anything that happened to him after the operation. Henry could repeat a telephone number with no difficulty, suggesting that his short-term memory store was just fine (Corkin, 2002, 2013; Hilts, 1995; Squire, 2009). But after information left the short-term store, it was gone forever. For example, he would often forget that he had just eaten a meal or fail to recognize the hospital staff who helped him on a daily basis. Studies of Henry and others have shown that the hippocampal region of the brain is critical for putting new information into the long-term store. When this region is damaged, individuals suffer from a condition known as anterograde amnesia, which is the inability to transfer new information from the short-term store into the long-term store.
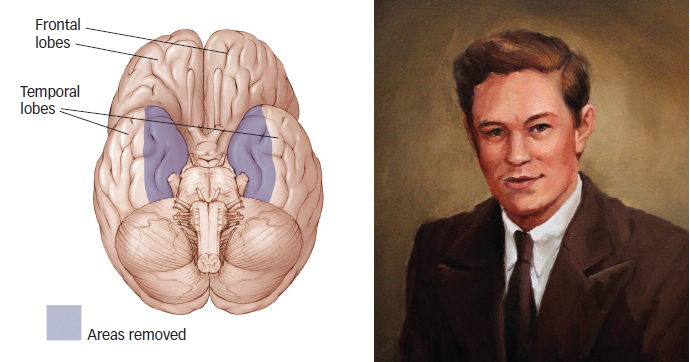
Figure 6.8: The Hippocampus Patient Henry Molaison had his hippocampus and adjacent structures of the medial temporal lobe (indicated by the shaded area) surgically removed to stop his epileptic seizures (left). As a result, he could not remember things that happened after the surgery. Henry (right), better known to the world as patient H.M., passed away on December 2, 2008, at the age of 82 at a nursing home near Hartford, Connecticut. Henry’s catastrophic inability to encode new long-term memories was discovered by Montreal psychologist Brenda Milner in 1953, and Molaison participated in countless memory experiments between that time and his death. In so doing, he made fundamental contributions to our understanding of memory and the brain.
Page 232
Some individuals with amnesia also suffer from retrograde amnesia, which is the inability to retrieve information that was acquired before a particular date, usually the date of an injury or surgery. The fact that Henry had much worse anterograde than retrograde amnesia suggests that the hippocampal region is not the site of long-term memory. Indeed, research has shown that different aspects of a single memory—its sights, sounds, smells, emotional content—are stored in different places in the cortex (Damasio, 1989; Schacter, 1996; Squire & Kandel, 1999). Some psychologists have argued that the hippocampal region acts as a kind of “index” that links together all of these otherwise separate bits and pieces so that we remember them as one memory (Schacter, 1996; Squire, 1992; Teyler & DiScenna, 1986). Over time, this index may become less necessary.
How is using the hippocampal region index like learning a recipe?
Going back to our cooking analogy, you can think of the hippocampal region index like a printed recipe. The first time you make a pie, you need the recipe to help you retrieve all the ingredients and then mix them together in the right amounts. As you bake more and more pies, though, you do not need to rely on the printed recipe anymore. Similarly, although the hippocampal region index is critical when a new memory is first formed, it may become less important as the memory ages. Another possibility is that the hippocampal index remains involved over long periods of time with some memories (highly detailed recollections of personal experiences, the kinds of memories that give us the feeling that we are almost reliving a past experience), but does not stay involved in less detailed, more general memories (Harand et al., 2012; Winocur, Moscovitch, & Bontempi, 2010). In terms of the cooking analogy, you might need to rely on a recipe each time you cook a complex meal with many details, but not when you cook a simpler meal with a less detailed recipe. Scientists are still debating the extent to which the hippocampal region helps us to remember details of our old memories (Bayley, Gold, et al., 2005; Kirwan et al., 2008; Moscovitch et al., 2006; Squire & Wixted, 2011; Winocur et al., 2010), but the notion of the hippocampus as an index explains why people like Henry cannot make new memories and why they can remember old ones.
6.2.3.2 Memory Consolidation
The idea that the hippocampus becomes less important over time for maintaining memories is closely related to the concept of consolidation, the process by which memories become stable in the brain (McGaugh, 2000). Shortly after encoding, memories exist in a fragile state in which they can be easily disrupted; once consolidation has occurred, they are more resistant to disruption. One type of consolidation operates over seconds or minutes. For example, when someone experiences a head injury in a car crash and later cannot recall what happened during the few seconds or minutes before the crash—but can recall other events normally—the head injury probably prevented consolidation of short-term memory into long-term memory. Another type of consolidation occurs over much longer periods of time—days, weeks, months, and years—and likely involves transfer of information from the hippocampus to more permanent storage sites in the cortex. The operation of this longer-term consolidation process is why individuals with retrograde amnesia with hippocampal damage can recall memories from childhood relatively normally, but are impaired when recalling experiences that occurred just a few years prior to the time they became amnesic (Kirwan et al., 2008; Squire & Wixted, 2011).
Page 233
How does a memory become consolidated? The act of recalling a memory, thinking about it, and talking about it with others probably contributes to consolidation (Moscovitch et al., 2006). And though you may not be aware of it, consolidation gets a boost from something that you do effortlessly every night: sleep. As explained in the Hot Science box, mounting evidence gathered during the past decade indicates that sleep plays an important role in memory consolidation.
HOT SCIENCE: Sleep on It
Thinking about pulling an all-nighter before your next big test? Here is a reason to reconsider: Our minds do not simply shut off when we sleep (see the Consciousness chapter), and in fact, sleep may be as important to our memories as wakefulness.
Nearly a century ago, Jenkins and Dallenbach (1924) reported that recall of recently learned information is greater immediately after sleeping than after the same amount of time spent awake. But Jenkins and Dallenbach did not think that sleep played an active role in strengthening or consolidating memory. They argued instead that being asleep passively protects us from encountering information that interferes with our ability to remember. As is explained by retroactive interference, that is a valid argument. However, during the past few years, evidence has accumulated that shows sleep plays an active role in memory consolidation, doing more than simply protecting us from waking interference (Diekelmann & Born, 2010; Ellenbogen, Payne, & Stickgold, 2006). Sleep selectively enhances the consolidation of memories that reflect the meaning or gist of an experience (Payne et al., 2009), as well as emotionally important memories (Payne et al., 2008), suggesting that sleep helps us to remember what is important and to discard what is trivial.
This idea is reinforced by recent evidence which shows that the beneficial effects of sleep on subsequent memory are observed only when people expect to be tested. In one study, Ines Wilhelm and her colleagues (2011) found that after studying a list of word pairs, participants who were informed that their memory would be tested later showed improved recall after sleep compared with an equivalent period of wakefulness. But a separate group that had not been informed of the memory test (and did not suspect it) showed no improvement in recall after sleep compared with wakefulness.
VAN DONGEN EV, THIELEN J-W, TAKASHIMA A, BARTH M, FERNÁNDEZ G (2012) SLEEP SUPPORTS SELECTIVE RETENTION OF ASSOCIATIVE MEMORIES BASED ON RELEVANCE FOR FUTURE UTILIZATION. PLOS ONE 7(8): E43426. DOI:10.1371/JOURNAL.PONE.0043426. © VAN DONGEN ET AL
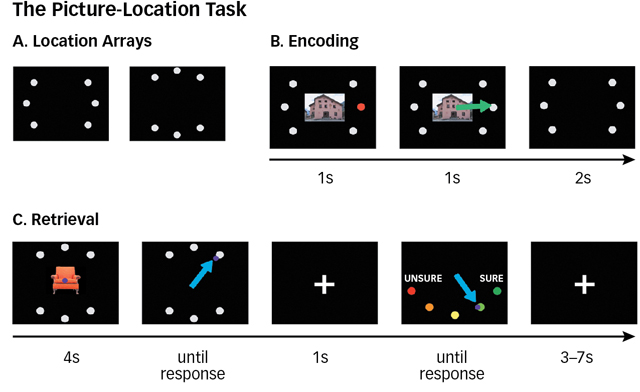
In an additional study (van Dongen et al., 2012), participants studied pictures of either buildings or furniture that were each associated with a particular location on a slide (see below). Soon after, participants received a memory test for the picture–location associations. They were then instructed that they would be retested 14 hours later for either the building pictures or the furniture pictures (the relevant category) and would not be retested on the other type of picture (the irrelevant category). Half of the participants were tested after sleep (initial learning was in the afternoon for these individuals) and half after remaining awake (initial learning was in the morning for these individuals). On the retest, there was less forgetting for the relevant than irrelevant categories in the sleep group, but there was no such difference in the wake group. Furthermore, for participants in the sleep group, the amount of time spent sleeping was correlated with retention of relevant information but not with retention of irrelevant information; those participants who slept longer retained more of the relevant information.
So, when you find yourself nodding off after hours of studying for your exam, the science is on the side of a good night’s sleep.
Page 234

Has seeing too many shark movies left you afraid to swim in the ocean? What evidence is there that someday we might be able to erase painful memories?
PM IMAGES/PHOTODISC/GETTY IMAGES
When is a consolidated memory vulnerable to disruption?
Many researchers have long believed that a fully consolidated memory becomes a permanent fixture in the brain, more difficult to get rid of than a computer virus. But another line of research that has developed rapidly in recent years suggests that things are not so simple. Experiments have shown that even seemingly consolidated memories can become vulnerable to disruption when they are recalled, thus requiring them to be consolidated again. This process is called reconsolidation (Dudai, 2012; Nader & Hardt, 2009). Evidence for reconsolidation mainly comes from experiments with rats showing that when animals are cued to retrieve a new memory that was acquired a day earlier, giving the animal a drug (or an electrical shock) that prevents initial consolidation will cause forgetting (Nader, Shafe, & LeDoux, 2000; Sara, 2000). Critically, if the animal is not actively retrieving the memory, the same drug (or shock) has no effect when given a day after initial encoding. This finding is surprising because it was once thought that when memories are consolidated, drugs or shock that prevent initial consolidation no longer have any impact. To the contrary, it appears that each time they are retrieved, memories become vulnerable to disruption and have to be reconsolidated.
Might it be possible one day to eliminate painful memories by disrupting reconsolidation? Recent research with traumatized individuals suggests it could be: When a traumatic event was reactivated after administration of a drug that reduces anxiety, there was a subsequent reduction in traumatic symptoms (Brunet et al., 2008, 2011). Other research with nontraumatized individuals has shown that something like this happens, without the use of drugs, as a result of reactivating a fear memory (being shocked in the presence of a particular object) a day after the memory was acquired. Adding nonfearful information to the reactivated memory (presenting the object again without shock) a few minutes later—when the memory is vulnerable to reconsolidation—resulted in long-lasting reduction of fear responses to the object, whereas adding nonfearful information to the reactivated memory 6 hours later—when the memory is no longer vulnerable to reconsolidation—did not have a long-lasting effect (Schiller et al., 2010). Related work indicates that disrupting reconsolidation can seemingly eliminate a conditioned fear memory in a part of the brain called the amygdala, which we will learn later in this chapter plays a key role in emotional memory (Agren et al., 2012). Reconsolidation thus appears to be a key memory process with many important implications.
6.2.4 Memories, Neurons, and Synapses

Nobel Prize–winning neuroscientist Eric Kandel took a risk and studied the tiny sea slug Aplysia based in part on a lesson he had learned from his wife regarding their recent marriage, which encouraged him to trust his intuition: “Denise was confident that our marriage would work, so I took a leap of faith and went ahead. I learned from that experience that there are many situations in which one cannot decide on the basis of cold facts alone—because the facts are often insufficient. One ultimately has to trust one’s unconscious, one’s instincts, one’s creative urge. I did this again in choosing Aplysia” (Kandel, 2006, p. 149).
SPENCER PLATT/NEWSMAKERS/GETTY IMAGES
We have already discussed parts of the brain that are related to memory storage, but we have not said much about how or where memories are stored. Research suggests that memory storage depends critically on the spaces between neurons. You will recall from the Neuroscience and Behaviour chapter that a synapse is the small space between the axon of one neuron and the dendrite of another, and neurons communicate by sending neurotransmitters across these synapses. As it turns out, sending a neurotransmitter across a synapse is not like sending a toy boat across a pond because the act of sending actually changes the synapse. Specifically, it strengthens the connection between the two neurons, making it easier for them to transmit to each other the next time. As the famous Canadian neuroscientist Donald Olding Hebb said, “cells that fire together wire together” (Hebb, 1949).
Page 235
The idea that the connections between neurons are strengthened by their communication, making communication easier the next time, provides the neurological basis for long-term memory, and much of what we know about this comes from the tiny sea slug Aplysia. The story of Aplysia and memory is closely linked with the work of neuroscientist Eric Kandel, who won the Nobel Prize in 2000 for his work with the creature. When Kandel first became interested in Aplysia back in the late 1950s, there were only two researchers in the entire world studying the tiny slug. But Aplysia was attractive to Kandel because it is relatively uncomplicated and has an extremely simple nervous system consisting of only 20 000 neurons (compared to roughly 100 billion in the human brain), so Kandel followed his intuition and studied Aplysia (Kandel, 2006).
When an experimenter stimulates Aplysia’s tail with a mild electric shock, the slug immediately withdraws its gill, and if the experimenter does it again a moment later, Aplysia withdraws its gill even more quickly. If the experimenter comes back an hour later and shocks Aplysia, the withdrawal of the gill happens as slowly as it did the first time, as if Aplysia cannot “remember” what happened an hour earlier (Abel et al., 1995). But if the experimenter shocks Aplysia over and over, it does develop an enduring “memory” that can last for days or even weeks. Research suggests that this long-term storage involves the growth of new synaptic connections between neurons (Abel et al., 1995; Kandel, 2006; Squire & Kandel, 1999). So, learning in Aplysia is based on changes involving the synapses for both short-term storage (enhanced neurotransmitter release) and long-term storage (growth of new synapses). Any experience that results in memory produces physical changes in the nervous system—even if you are a slug.

By studying the sea slug Aplysia californica’s extremely simple nervous system, researchers were able to determine that long-term memory storage depends on the growth of new synaptic connections between neurons.
© DONNA IKENBERRY/ART DIRECTORS & TRIP/ALAMY
How does building a memory produce a physical change in the nervous system?
If you are something more complex than a slug—say, a chimpanzee or your roommate—a similar process of synaptic strengthening happens in the hippocampus, which we have seen is an area crucial for storing new long-term memories. In the early 1970s, researchers applied a brief electrical stimulus to a neural pathway in a rat’s hippocampus (Bliss & Lømo, 1973). They found that the electrical current produced a stronger connection between synapses that lay along the pathway and that the strengthening lasted for hours or even weeks. They called this long-term potentiation (more commonly known as LTP), a process whereby communication across the synapse between neurons strengthens the connection, making further communication easier. Long-term potentiation has a number of properties that indicate to researchers that it plays an important role in long-term memory storage: It occurs in several pathways within the hippocampus; it can be induced rapidly; and it can last for a long time. In fact, drugs that block LTP can turn rats into rodent versions of Henry Molaison: The animals have great difficulty remembering where they have been recently and become easily lost in a maze (Bliss, 1999; Morris et al., 1986).
Page 236
There are several different types of memory storage:
Sensory memory holds information for a second or two.
Short-term or working memory retains information for about 15 to 20 seconds.
Long-term memory stores information anywhere from minutes to years or decades.
The hippocampus and nearby structures play an important role in long-term memory storage, as shown by the severe amnesia of individuals such as Henry Molaison. The hippocampus is also important for memory consolidation, the process that makes memories increasingly resistant to disruption over time. Sleep contributes importantly to memory consolidation.
Memory storage depends on changes in synapses, and long-term potentiation (LTP) increases synaptic connections.