27.3Ketone Bodies Are Another Fuel Source Derived from Fats
✓ 2 Describe ketone bodies and their role in metabolism.
Most acetyl CoA produced by fatty acid degradation enters the citric acid cycle. However, some acetyl CoA units are used to form an alternative fuel source called ketone bodies—namely, acetoacetate, d-3-hydroxybutyrate (β-hydroxybutyrate), and acetone. Although ketone bodies do not generate as much ATP as do the fatty acids from which they are derived, they have the advantage of being water soluble, and so they are an easily transportable form of acetyl units. Originally thought to be indicators of impaired metabolism, acetoacetate and d-3-hydroxybutyrate are now known to be normal fuels of respiration. Indeed, heart muscle and the renal cortex of the kidney may use acetoacetate in preference to glucose. Acetone is produced by the slow, spontaneous decarboxylation of acetoacetate. Under starvation conditions, acetone may be captured to synthesize glucose.
Ketone-Body Synthesis Takes Place in the Liver
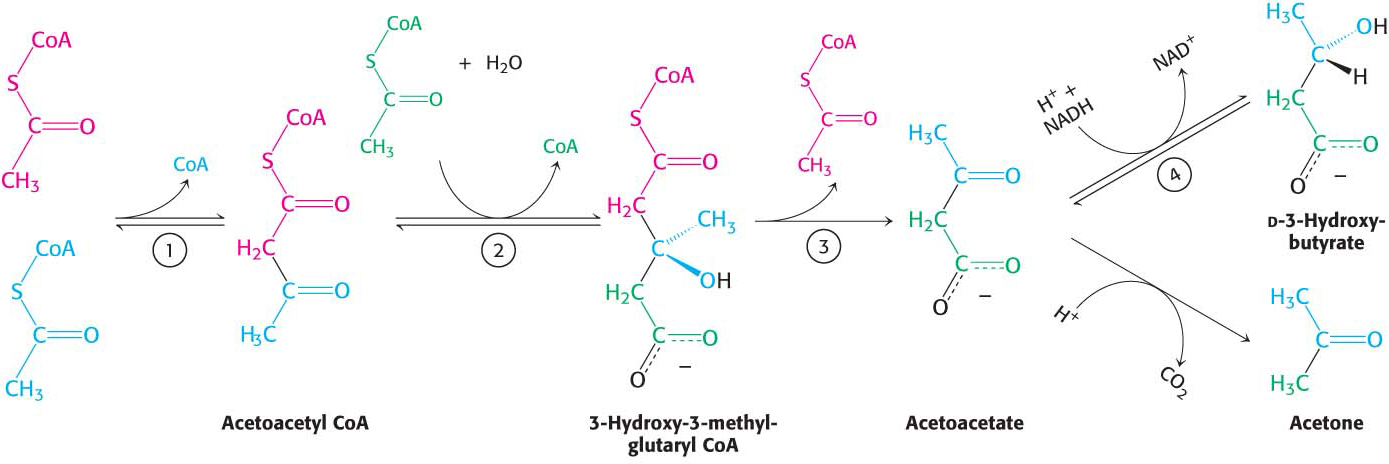
The major site of ketogenesis—the production of acetoacetate and d-3-hydroxybutyrate—is the mitochondria of the liver. Acetoacetate is formed from acetyl CoA in three steps (Figure 27.10). The sum of these reactions is
Figure 27.10 The formation of ketone bodies. The ketone bodies—acetoacetate, d-3-hydroxybutyrate, and acetone—are formed from acetyl CoA primarily in the liver. Enzymes catalyzing these reactions are (1) 3-ketothiolase, (2) hydroxymethylglutaryl CoA synthase, (3) hydroxymethylglutaryl CoA cleavage enzyme, and (4) d-3-hydroxybutyrate dehydrogenase. Acetoacetate spontaneously decarboxylates to form acetone.
?
QUICK QUIZ 2
Why might d-3-hydroxybutyrate be considered a superior ketone body compared with acetoacetate?
d-3-Hydroxybutyrate is more energy rich because its oxidation potential is greater than that of acetoacetate. After having been absorbed by a cell, d-3-hydroxybutyrate is oxidized to acetoacetate, generating high-energy electrons in the form of NADH. The acetoacetate is then cleaved to yield to acetyl CoA.
A fourth step is required to form d-3-hydroxybutyrate: the reduction of acetoacetate in the mitochondrial matrix. The ketone bodies then exit the liver mitochondria into the blood, with the assistance of specific carrier proteins, and are transported to peripheral tissues.
Page 498
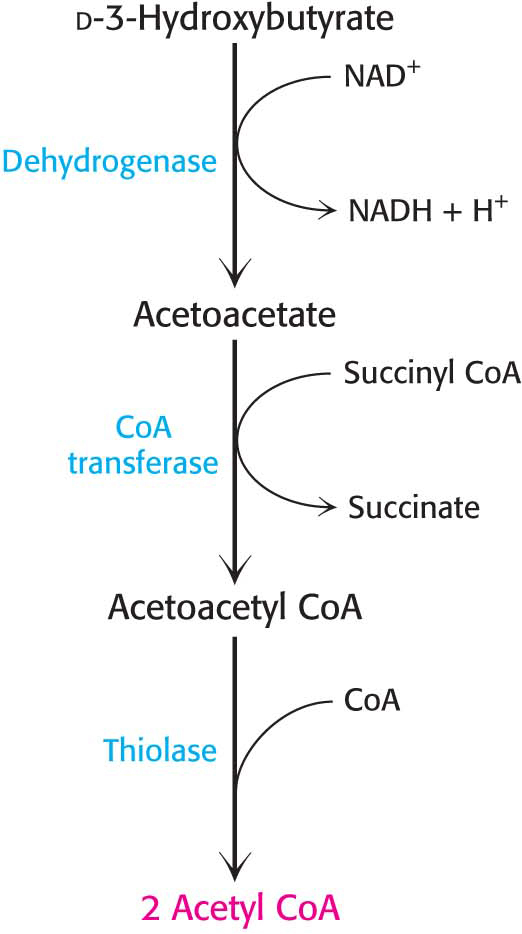
Figure 27.11 The utilization of d-3-hydroxybutyrate and acetoacetate as a fuel. d-3-Hydroxybutyrate is oxidized to acetoacetate with the formation of NADH. Acetoacetate is then converted into two molecules of acetyl CoA, which then enter the citric acid cycle.
How are ketone bodies used as a fuel? Figure 27.11 shows how ketone bodies are metabolized to generate NADH as well as acetyl CoA, the fuel for the citric acid cycle. d-3-Hydroxybutyrate is oxidized to acetoacetate, which is then activated by a specific CoA transferase. Finally, acetoacetyl CoA is cleaved by a thiolase to yield two molecules of acetyl CoA, which enter the citric acid cycle. Liver cells, however, lack the CoA transferase, thus allowing acetoacetate to escape the liver and be transported out of the cell and to other tissues, rather than being metabolized in the liver. Acetoacetate also has a regulatory role. High levels of acetoacetate in the blood signify an abundance of acetyl units and lead to a decrease in the rate of lipolysis in adipose tissue.
CLINICAL INSIGHT
Ketogenic Diets May Have Therapeutic Properties
Interestingly, diets that promote ketone-body formation, called ketogenic diets, are frequently used as a therapeutic option for children with drug-resistant epilepsy. Ketogenic diets are rich in fats and low in carbohydrates, with adequate amounts of protein. In essence, the body is forced into starvation mode, where fats and ketone bodies become the main fuel source. How such diets reduce the seizures suffered by the children is currently unknown.
Animals Cannot Convert Fatty Acids into Glucose
A typical human being has far greater fat stores than glycogen stores. However, glycogen is necessary to fuel very active muscle as well as the brain, which normally uses only glucose as a fuel. When glycogen levels are low, why can’t the body make use of fat stores instead and convert fatty acids into glucose? Because animals are unable to effect the net synthesis of glucose from fatty acids. Specifically, the acetyl CoA generated by fatty acid degradation cannot be converted into pyruvate or oxaloacetate in animals. Recall that the reaction that generates acetyl CoA from pyruvate is irreversible. The two carbon atoms of the acetyl group of acetyl CoA enter the citric acid cycle, but two carbon atoms leave the cycle in decarboxylations catalyzed by isocitrate dehydrogenase and α-ketoglutarate dehydrogenase. Consequently, oxaloacetate is regenerated, but it is not formed de novo when the acetyl unit of acetyl CoA is oxidized by the citric acid cycle. In essence, two carbon atoms enter the cycle as an acetyl group, but two carbons leave the cycle as CO2 before oxaloacetate is generated. Consequently, no net synthesis of oxaloacetate is possible. In contrast, plants have two additional enzymes enabling them to convert the carbon atoms of acetyl CoA into oxaloacetate.

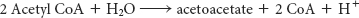
 CLINICAL INSIGHT
CLINICAL INSIGHT