2.2 The Chromosomal Basis of Single-Gene Inheritance Patterns
Mendel’s view of equal segregation was that the members of a gene pair segregated equally in gamete formation. He did not know about the subcellular events that take place when cells divide in the course of gamete formation. Now we understand that gene pairs are located on chromosome pairs and that it is the members of a chromosome pair that actually segregate, carrying the genes with them. The members of a gene pair are segregated as an inevitable consequence.
Single-gene inheritance in diploids
When cells divide, so must the nucleus and its main contents, the chromosomes. To understand gene segregation, we must first understand and contrast the two types of nuclear divisions that take place in eukaryotic cells. When somatic (body) cells divide to increase their number, the accompanying nuclear division is called mitosis, a programmed stage of all eukaryotic cell-
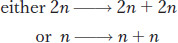
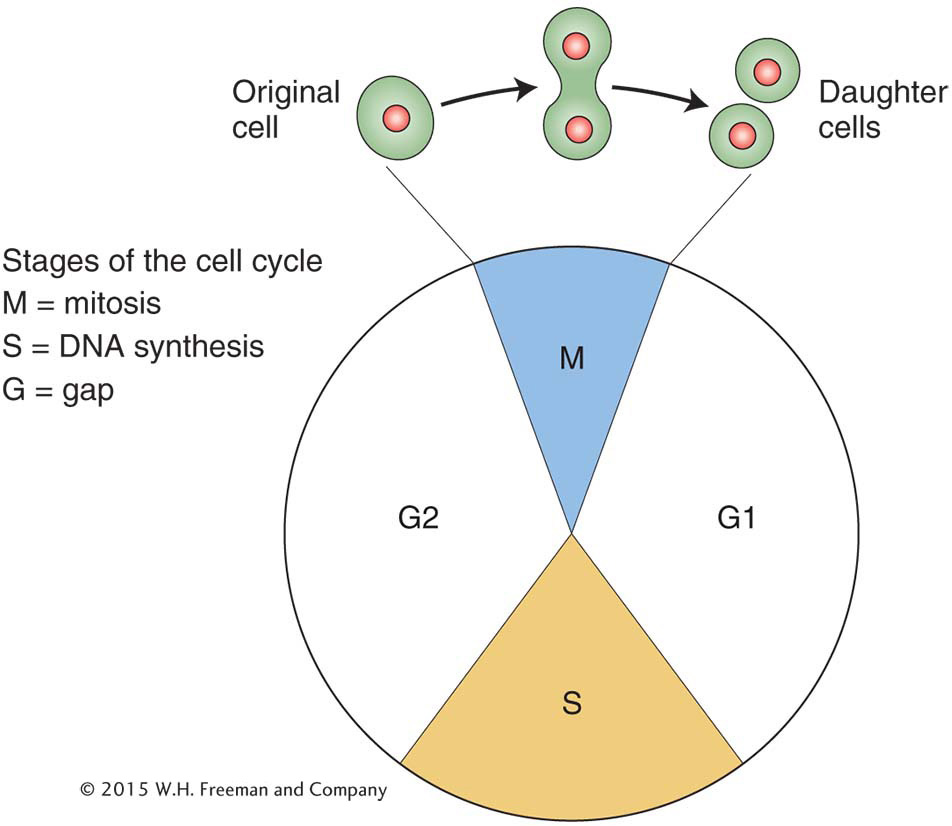
This “trick” of constancy is accomplished when each chromosome replicates to make two identical copies of itself, with underlying DNA replication. The two identical copies, which are often visually discernible, are called sister chromatids. Then, each copy is pulled to opposite ends of the cell. When the cell divides, each daughter cell has the same chromosomal set as its progenitor.
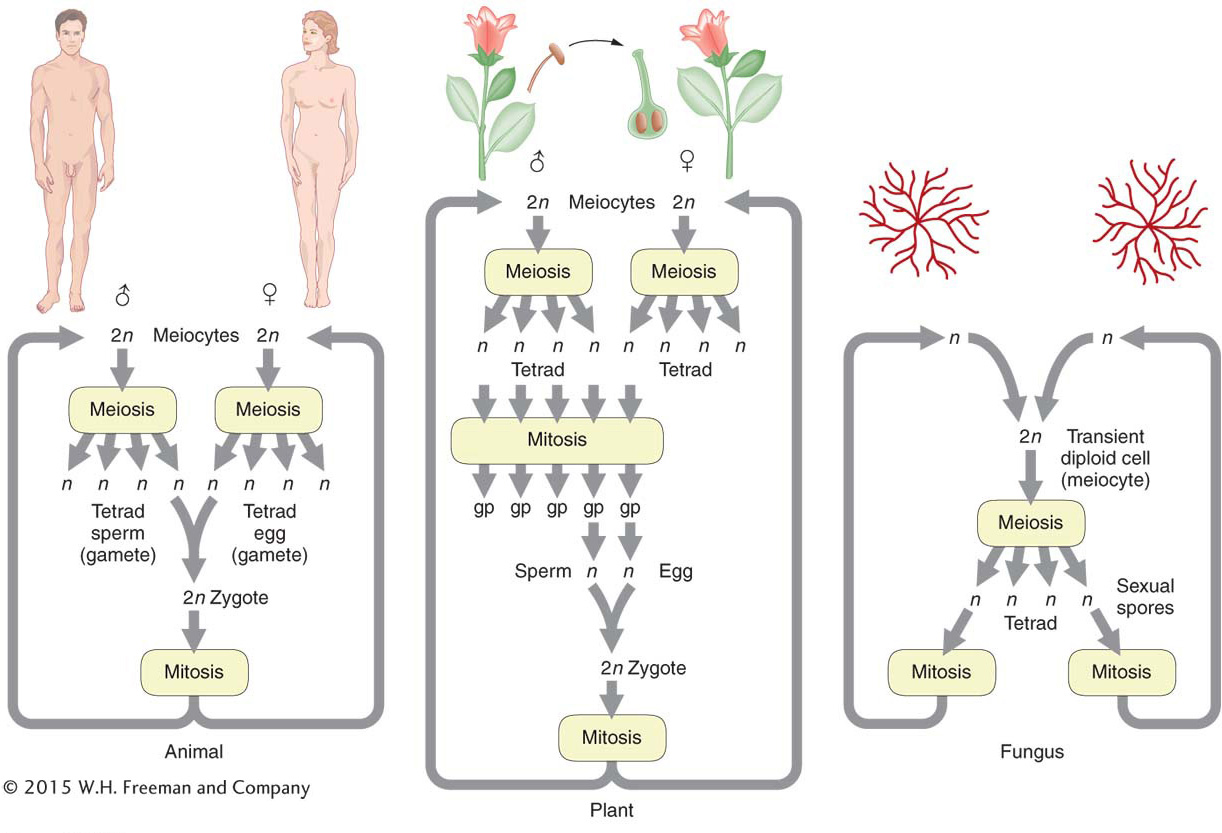
In addition, most eukaryotes have a sexual cycle, and, in these organisms, specialized diploid cells called meiocytes are set aside that divide to produce sex cells such as sperm and egg in plants and animals, or sexual spores in fungi or algae. Two sequential cell divisions take place, and the two nuclear divisions that accompany them are called meiosis. Because there are two divisions, four cells are produced from each progenitor cell. Meiosis takes place only in diploid cells, and the resulting gametes (sperm and eggs in animals and plants) are haploid. Hence, the net result of meiosis is
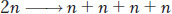
This overall halving of chromosome number during meiosis is achieved through one replication and two divisions. As with mitosis, each chromosome replicates once, but in meiosis the replicated chromosomes (sister chromatids) remain attached. One of each of the replicated chromosome pairs is pulled to opposite ends of the cell, and division occurs. At the second division, the sister chromatids separate and are pulled to opposite ends of the cell.
The location of the meiocytes in animal, plant, and fungal life cycles is shown in Figure 2-7.
The basic genetic features of mitosis and meiosis are summarized in Figure 2-8. To make comparison easier, both processes are shown in a diploid cell. Notice, again, that mitosis takes place in one cell division, and the two resulting “daughter” cells have the same genomic content as that of the “mother” (progenitor) cell. The first key process to note is a premitotic chromosome replication. At the DNA level, this stage is the synthesis, or S, phase (see Figure 2-6), at which the DNA is replicated. The replication produces pairs of identical sister chromatids, which become visible at the beginning of mitosis. When a cell divides, each member of a pair of sister chromatids is pulled into a daughter cell, where it assumes the role of a fully fledged chromosome. Hence, each daughter cell has the same chromosomal content as the original cell.
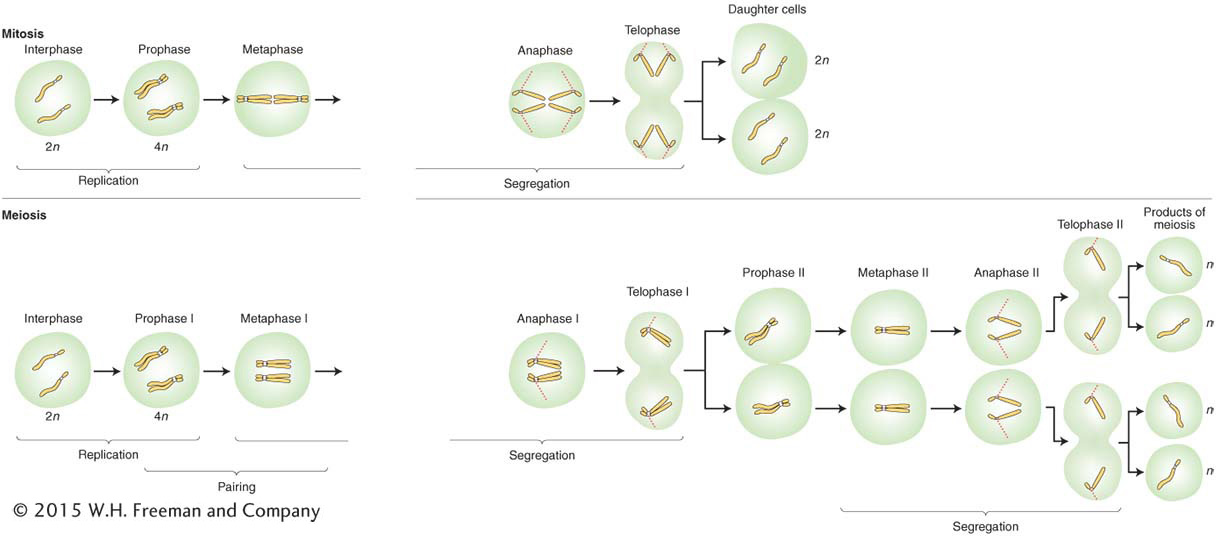
Before meiosis, as in mitosis, chromosome replication takes place to form sister chromatids, which become visible at meiosis. The centromere appears not to divide at this stage, whereas it does in mitosis. Also in contrast with mitosis, the homologous pairs of sister chromatids now unite to form a bundle of four homologous chromatids. This joining of the homologous pairs is called synapsis, and it relies on the properties of a macromolecular assemblage called the synaptonemal complex (SC), which runs down the center of the pair. Replicate sister chromosomes are together called a dyad (from the Greek word for “two”). The unit comprising the pair of synapsed dyads is called a bivalent. The four chromatids that make up a bivalent are called a tetrad (Greek for “four”), to indicate that there are four homologous units in the bundle.
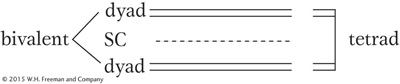
(A parenthetical note. The process of crossing over takes place at this tetrad stage. Crossing over changes the combinations of alleles of several different genes but does not directly affect single-
The bivalents of all chromosomes move to the cell’s equator, and, when the cell divides, one dyad moves into each new cell, pulled by spindle fibers attached near the centromeres. In the second cell division of meiosis, the centromeres divide and each member of a dyad (each member of a pair of chromatids) moves into a daughter cell. Hence, although the process starts with the same genomic content as that for mitosis, the two successive segregations result in four haploid cells. Each of the four haploid cells that constitute the four products of meiosis contains one member of a tetrad; hence, the group of four cells is sometimes called a tetrad, too. Meiosis can be summarized as follows:
Start: → two homologs
Replication: → two dyads
Pairing: → tetrad
First division: → one dyad to each daughter cell
Second division: → one chromatid to each daughter cell
Research in cell biology has shown that the spindle fibers that pull apart chromosomes are polymers of the molecule tubulin. The pulling apart is caused mainly by a depolymerization and hence shortening of the fibers at the point where they are attached to the chromosomes.
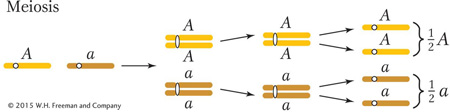
The behavior of chromosomes during meiosis clearly explains Mendel’s law of equal segregation. Consider a heterozygote of general type A/a. We can simply follow the preceding summary while considering what happens to the alleles of this gene:
Start: one homolog carries A and one carries a
Replication: one dyad is AA and one is aa
Pairing: tetrad is A/A/a/a
First-
Second-
Hence, the products of meiosis from a heterozygous meiocyte A/a are  A and
a, precisely the equal ratio that is needed to explain Mendel’s first law.
A and
a, precisely the equal ratio that is needed to explain Mendel’s first law.
Note that we have focused on the broad genetic aspects of meiosis necessary to explain single-
Single-gene inheritance in haploids
We have seen that the cellular basis of the law of equal segregation is the segregation of chromosomes in the first division of meiosis. In the discussion so far, the evidence for the equal segregation of alleles in meiocytes of both plants and animals is indirect, based on the observation that crosses show the appropriate ratios of progeny expected under equal segregation. Recognize that the gametes in these studies (such as Mendel’s) must have come from many different meiocytes. However, in some organisms, their special life cycle allows the examination of the products of one single meiocyte. These organisms are called haploids, of which good examples are most fungi and algae. They spend most of their lives in the haploid state but can mate, in the process forming a transient diploid cell that becomes the meiocyte. In some species, the four products of a single meiosis are temporarily held together in a type of sac.
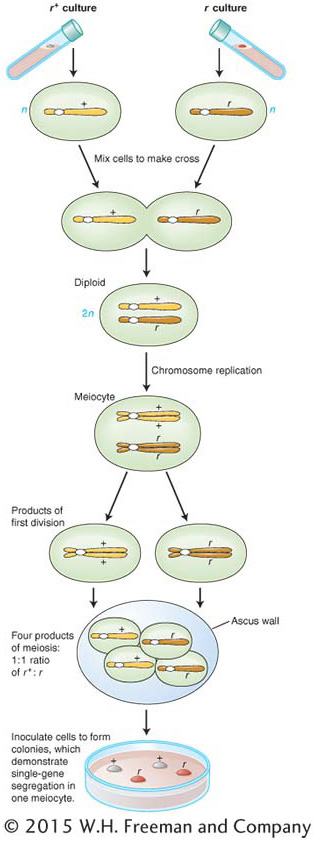
Baker’s yeast, Saccharomyces cerevisiae (a fungus), provides a good example (see the yeast Model Organism box in Chapter 12). In fungi, there are simple forms of sexes called mating types. In S. cerevisiae, there are two mating types, and a successful cross can only occur between strains of different mating types. Let’s look at a cross that includes a yeast mutant. Normal wild-
r+ × r
When two cells of opposite mating type fuse, a diploid cell is formed, and it is this cell that becomes the meiocyte. In the present example, the diploid meiocyte would be heterozygous, r+/r. Replication and segregation of r+ and r would give a tetrad of two meiotic products (spores) of genotype r+ and two of r, all contained within a membranous sac called an ascus. Hence,
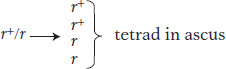
The details of the process are shown in Figure 2-9. If the four spores from one ascus are isolated (representing a tetrad of chromatids) and used to generate four yeast cultures, then equal segregation within one meiocyte is revealed directly as two white cultures and two red. If we analyzed the random spores from many meiocytes, we would find about 50 percent red and 50 percent white.
Note the simplicity of haploid genetics: a cross requires the analysis of only one meiosis; in contrast, a diploid cross requires a consideration of meiosis in both the male and the female parent. This simplicity is an important reason for using haploids as model organisms. Another reason is that, in haploids, all alleles are expressed in the phenotype because there is no masking of recessives by dominant alleles on the other homolog.