15.5 Regulation of Transposable Element Movement by the Host
Genetic analysis is a very powerful tool used to dissect complex biological processes through the isolation of mutants and, ultimately, mutant genes (for example, see Chapter 13 on development). Many laboratories around the world are using genetic analyses to identify the host genes responsible for repressing the movement of transposable elements and, in this way, maintaining the stability of the genome.
The repression of transposable elements was first investigated in the laboratory of Ron Plasterk in the late 1990s, using the model organism C. elegans (a nematode; see the Model Organism box in Chapter 13). This story starts with the observation of a striking difference between the mobility of a transposable element called Tc1 in two different cell types of the model organism. Tc1 is a DNA transposon that, like the Ac element of maize, can lead to an unstable mutant phenotype when it excises from a gene with a visible phenotype (see Figure 15-4). There are 32 Tc1 elements in the sequenced genome of the common laboratory strain called Bristol. Significantly, Tc1 transposes in somatic but not germ-line cells. That observation suggested to Plasterk that transposition is repressed in the germ line by the host. Evidently, germ-line repression results from the silencing of the transposase genes of all 32 Tc1 copies in germ-line cells. Can you propose an explanation for why it makes biological sense for a host to repress transposition in the germ line and not in the soma?
Page 572
Figure 15-25: A mutant search leads to the genes required to repress transposition
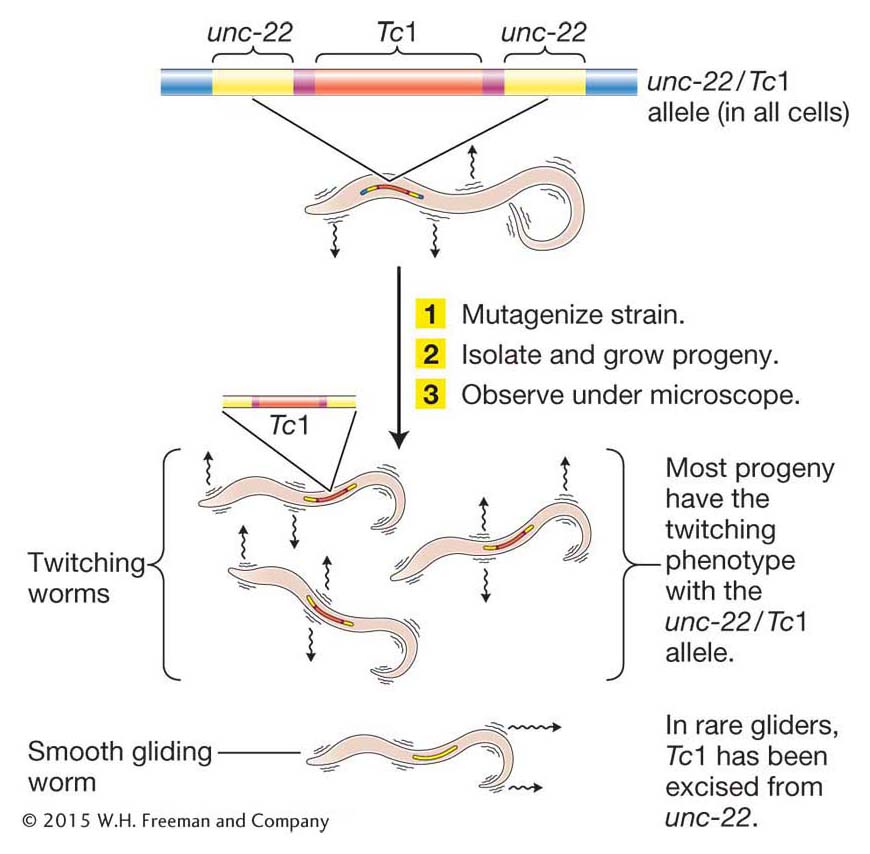
Figure 15-25: Experimental design used to identify genes required to repress transposition. Investigators look for mutants that have regained normal movement because mutations in these individuals would have disabled the repression mechanism that prevents the transposition of the Tc 1 element from the unc-22 gene.
Plasterk and his co-workers set out to identify C. elegans genes responsible for silencing the transposase gene. They began with a C. elegans strain that had Tc1 inserted in the unc-22 gene (designated unc-22/Tc1; Figure 15-25). This was the same gene that was silenced in the experiment of Fire and Mello that led to their sharing the Nobel Prize (see Chapter 8). Whereas wild-type C. elegans glides smoothly on the surface of the agar in a petri dish (as illustrated by horizontal arrows in Figure 15-25), worms with the mutant unc-22/Tc1 gene have a twitching movement (as illustrated by vertical arrows in Figure 15-25) that can be easily observed with a microscope. Because Tc1 cannot normally transpose in the germ line, it remains inserted in the unc-22 gene and continues to disrupt its function. Thus, the strain with the mutant unc-22/Tc1 gene should express a twitching phenotype from generation to generation. However, Plasterk and co-workers reasoned that mutations that inactivated C. elegans genes required for repression would allow Tc1 to excise from the unc-22/Tc1 allele in the germ line and revert the twitching phenotype to wild type (unc-22). To this end, they exposed the mutant unc-22/Tc1 strain to a chemical that greatly increased the frequency of mutation (called a mutagen; see Chapter 16) and examined their progeny under a microscope, searching for rare worms that no longer twitched.
This and subsequent genetic screens identified over 25 C. elegans genes that, when mutated, allowed the host to excise Tc1 in the germ line. Significantly, many of the products of these genes are integral components of the RNAi silencing pathway, including proteins found in Dicer and RISC (see Chapters 8 and 12). Recall from Chapter 8 that Dicer binds to long dsRNAs and cleaves them into small dsRNA fragments. These fragments are then unwound so that one strand, the siRNA, can target RISC to chop up complementary mRNAs (see Figure 8-24).
Beginning with this elegant genetic screen, many years of experimentation have led to the following model for the repression of transposable elements in the germ line of C. elegans. With 32 Tc 1 elements scattered throughout the C. elegans genome, a few elements near genes are transcribed along with the nearby gene by “read-through” transcription (see Chapter 8). Because the ends of Tc 1 are 54-bp terminal inverted repeats, the Tc1 RNA spontaneously forms dsRNA (Figure 15-26). Like all dsRNAs produced in most eukaryotes, this RNA is recognized by Dicer and ultimately siRNA is produced, which directs RISC to chop up complementary Tc1 transcripts. Because all Tc1 RNA is efficiently chopped up in the germ line, the element-encoded transposase gene is silenced. Without transposase, the element cannot excise. It has been hypothesized that Tc1 can transpose in somatic cells because RNAi is not as efficient and some transposase can be produced.
Figure 15-26: A single Tc 1 element can repress transposition
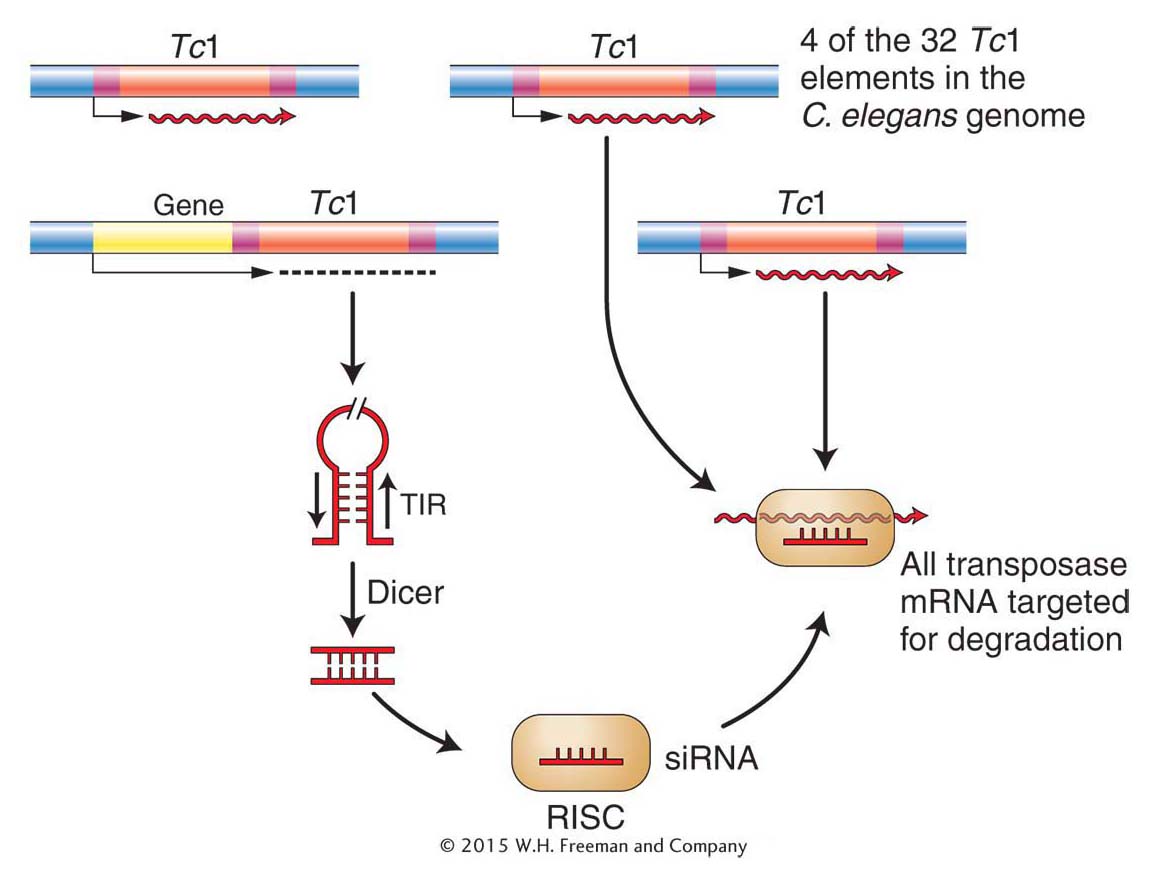
Figure 15-26: The production of dsRNA from only a single Tc1 element is sufficient to silence all of the Tc1 transposase genes and thereby repress transposition in the germ line. The siRNA derived from Tc 1 dsRNA is bound to RISC and targets all complementary RNA for degradation.
Over the past decade, numerous laboratories working with both plants and animals have discovered that mutations that disrupt the RNAi pathway often lead to the activation of transposable elements in their respective genomes. Because of the abundance of transposable elements in eukaryotic genomes and their ancient association, it has been suggested that the natural function of the RNAi pathway is to maintain genome stability by repressing the movement of transposable elements.
Page 573
KEY CONCEPT
Eukaryotic hosts use RNAi to repress the expression of active transposable elements in their genomes. In this way, a single element that inserts near a gene can be transcribed to produce dsRNA that will trigger the silencing of all copies of the element in the genome.
Genome surveillance in animals and bacteria
The RNAi silencing pathway is akin to radar in that the host is able to detect new insertions of transposons into the genome if they generate antisense RNA. The host then responds by producing siRNAs that target transposase mRNA, silencing the gene and preventing the movement of all transposable-element (TE) family members. Recently two other types of genome radar (also called genome surveillance) have been described that utilize different classes of small noncoding RNAs to target “invasive” nucleic acids including transposons and viruses (see Chapter 5). Although not yet completely understood, these mechanisms are presented here because they illustrate how different solutions evolve to solve similar biological problems.
piRNAs in animals In the germ lines of several animal species including Drosophila and mammals, active transposons are repressed through the action of piRNAs (short for Piwi-interacting RNAs). Like siRNAs, piRNAs are short single-stranded RNAs (26–30 nt in mammals) that interact with a protein complex (in this case, one that contains the protein piwi-Argonaute from which it derives its name). Once associated, piRNAs guide piwi-Argonaute to degrade complementary mRNAs (Figure 15-27). Unlike siRNAs, piRNAs do not originate from the double-stranded RNA pathway shown in Figure 15-26 for Tc1. Instead, and quite ingeniously, animal genomes contain several long (often >100 kb) loci called pi-clusters that serve as traps to ensnare active transposons. A pi-cluster is comprised of remnants of many different transposons that represent a historical record of prior insertions of active transposons into that locus.
Figure 15-27: Inactivation of TEs following insertion into pi-clusters
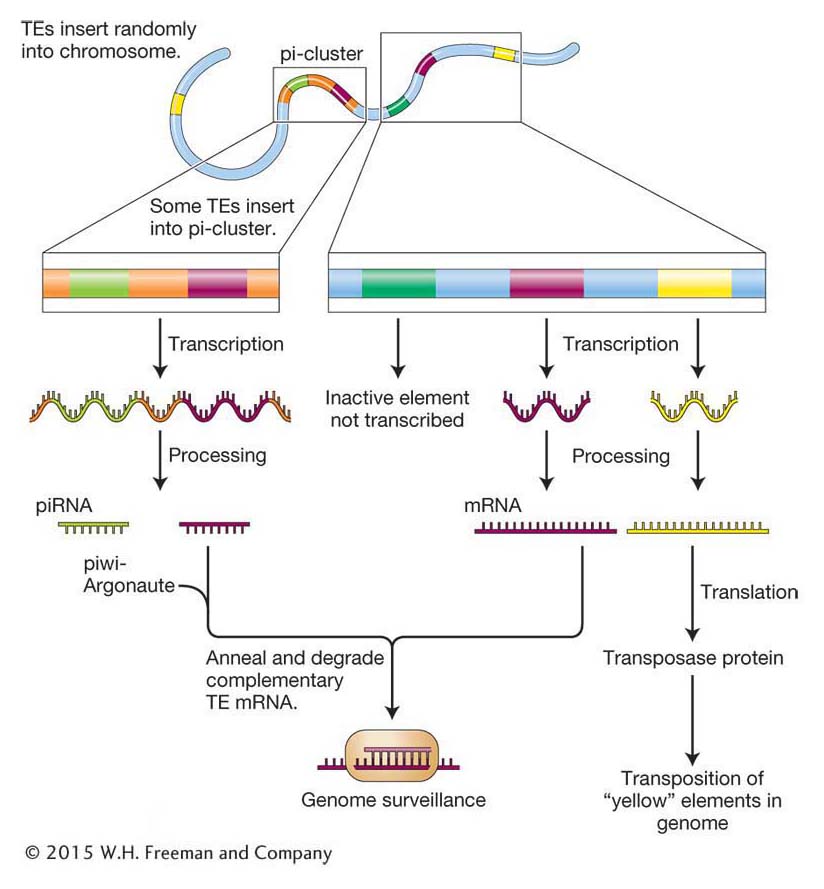
Figure 15-27: Insertion of the green and pink transposons into a pi-cluster in the genome results in the degradation of transcripts from these two transposons by the steps shown and described in the text. In contrast, the yellow transposon will remain active until copies insert by chance into a pi-cluster.
Figure 15-28: Acquisition of DNA by the CRISPR locus in some bacterial species
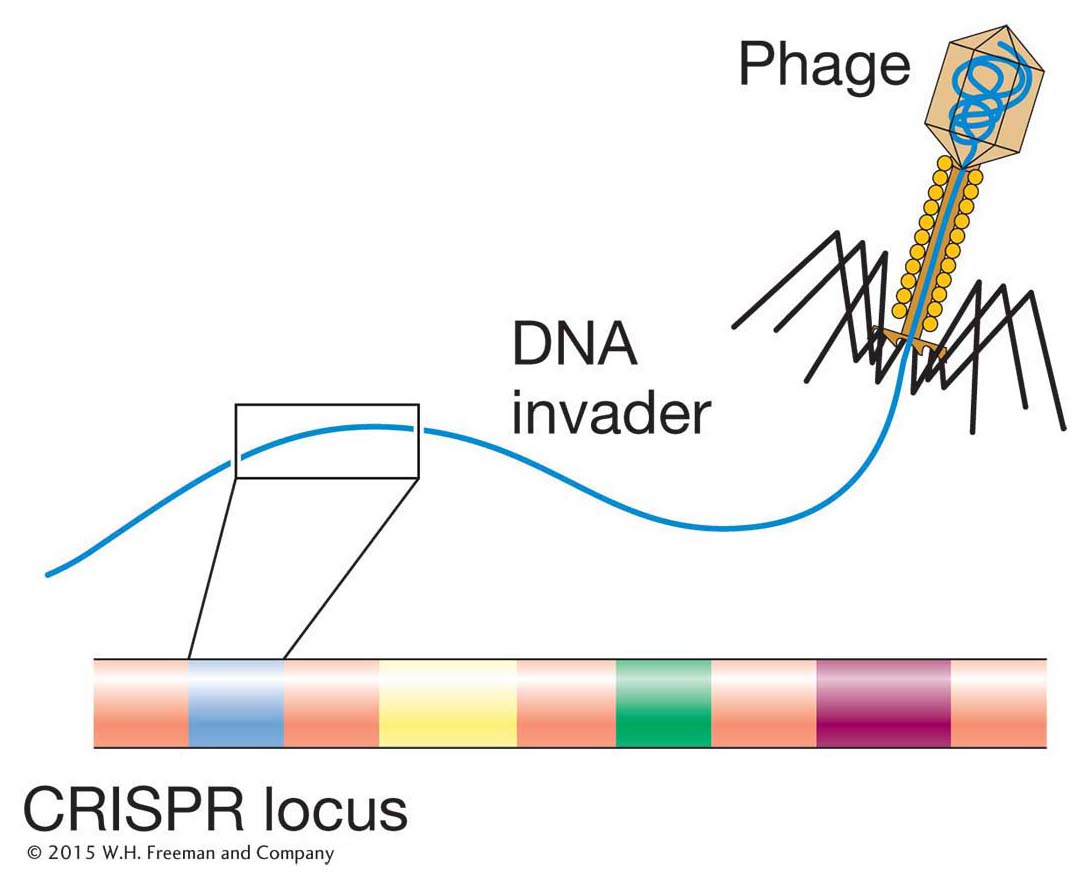
Figure 15-28: Acquisition of DNA by the CRISPR locus in some bacterial species. Part of the DNA from an invading phage genome (shown in yellow) is incorporated into the CRISPR locus by an unknown mechanism.
The first step in host genome surveillance is the insertion of a transposon into one of several pi-cluster loci scattered around the genome. Transcription of pi-clusters produce long RNAs that may include antisense RNA from the newly inserted element. These long RNAs are then processed into the final piRNAs that associate with piwi-Argonaute and go on to degrade transposon-derived mRNAs transcribed from anywhere in the genome. Thus, an active TE inserting randomly throughout the genome is recognized by genome surveillance only when it happens to insert into a pi-cluster and become a permanent part of the locus.
Page 574
crRNAs in bacteria Nucleic acid molecules usually invade bacterial species during bacteriophage infection (see Chapter 5) when viral DNA genomes are injected into bacteria (see Figure 5-22). In an antiviral pathway that is still being elucidated, fragments of the invading virus genome are captured by bacterial loci called CRISPR (clustered regularly interspaced short palindromic repeats) (Figure 15-28), where they are transcribed into long RNAs that are processed into short crRNAs. Much like siRNAs and piRNAs, crRNAs interact with and guide bacterial protein complexes to degrade complementary RNAs from the invading viral genome.
One shared feature of the pi-clusters and CRISPR loci is that new insertions of transposon or viral DNA fragments, respectively, result in a permanent, genetic change in these loci that is inherited by their progeny.
KEY CONCEPT
Like siRNAs, piRNAs in animals and crRNAs in bacteria interact with protein complexes and guide them to degrade complementary sequences in transposons and viruses, respectively. These small noncoding RNAs have their origin in long RNAs transcribed from loci that capture fragments of invasive DNA.Page 575
WHAT GENETICISTS ARE DOING TODAY
Much like planes that evade radar by flying close to the ground, some transposons have evolved mechanisms that allow them to evade the RNAi silencing pathway. These transposons can attain very high copy numbers. Evidence for these mechanisms can be found in the genomes of all characterized plants and animals containing transposon families (such as Alu in humans) that have thousands of members.
How do some transposons avoid detection by the RNAi silencing pathway? The short answer is that in most cases, we do not know. To understand how a transposon avoids detection, it is necessary to study actively transposing elements. To date, scientists have detected very few transposon families with high copy numbers that are still actively transposing. One of the best-characterized elements among this small group is a special type of nonautonomous DNA transposon called a miniature inverted repeat transposable element (abbreviated MITE). Like other nonautonomous elements, MITEs can form by deletion of the transposase gene from an autonomous element. However, unlike most nonautonomous elements, MITEs can attain very high copy numbers, particularly in the genomes of some grasses (see Figure 15-23). Some MITEs in grasses have been amplified to thousands of copies.
The only actively transposing MITE isolated to date is the mPing element of rice, which is formed from the autonomous Ping element by deletion of the entire transposase gene (Figure 15-29). This element was discovered in the laboratory of Susan Wessler by Ning Jiang. Another member of the Wessler laboratory, Ken Naito, documented that in individuals of some rice strains there are only 3 to 7 copies of Ping and over 1000 copies of mPing. Remarkably, the copy number of mPing in these strains is increasing by almost 40 new insertions per plant per generation.
Figure 15-29: MITEs attain very high copy numbers
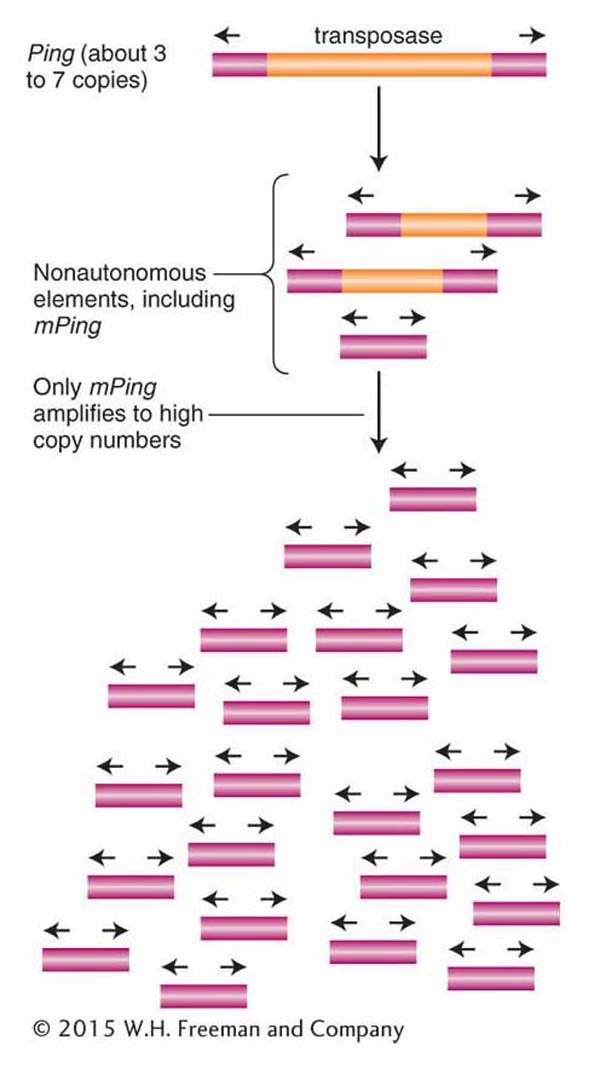
Figure 15-29: MITEs are nonautonomous DNA transposons that can attain very high copy numbers because they do not encode the transposase necessary for their transposition. The active MITE mPing is the only deletion derivative of the autonomous Ping element that has attained high copy number in certain strains of rice.
Page 576
Two questions about the rapid increase in mPing copy number immediately come to mind. First, how does a rice strain survive a transposable-element burst of this magnitude? To address this question, the Wessler laboratory used next-generation sequencing technology (see Chapter 14) to determine the insertion sites of over 1700 mPing elements in the rice genome. Surprisingly, they found that the element avoided inserting into exons, thus minimizing the impact of insertion on rice gene expression. The mechanism underlying this preference is currently being investigated.
The second question is, why does the rice host apparently fail to repress mPing transposition? While this question is also an active area of current research, a reasonable hypothesis is that mPing can fly under the hosts’ RNAi radar because it does not contain any part of the transposase gene that resides on the Ping element (Figure 15-29). Thus, read-through transcription into mPing elements inserted throughout the rice genome will produce lots of dsRNA and siRNA. However, because siRNAs derived from mPing share no sequence with the source of transposase, siRNAs produced from mPing will not induce silencing mechanisms aimed at transposase. Instead, the transposase gene will remain active and will continue to catalyze the movement of mPing. According to this hypothesis, mPing transposition will be repressed only when a much rarer Ping insertion generates dsRNA that triggers the silencing of its transposase gene.
KEY CONCEPT
MITEs are nonautonomous DNA transposons that can attain high copy numbers. While MITEs can utilize the transposase of autonomous elements, they probably evade host repression because their amplification does not lead to the silencing of the transposase gene.