12-3 Evolution, Environment, and Behavior
Odor and taste play a fundamental role in the biology of emotional and motivated behavior. Why does the sight or smell of a bird or a mouse trigger stalking and killing in a cat? Why does the human body stimulate sexual interest? We can address such questions by investigating the evolutionary and environmental influences on brain circuit activity that contribute to behavior.
Evolutionary Influences on Behavior
One evolutionary explanation hinges on the concept of innate releasing mechanisms (IRMs), activators for inborn adaptive responses that aid an animal’s survival. IRMs help an animal to feed, reproduce, and escape predators. The concept is best understood by analyzing its parts.
IRMs are present from birth rather than acquired through experience, as the term innate implies. These mechanisms have proved adaptive and therefore have been maintained in the genome of the species. The term releasing indicates that IRMs act as triggers for behaviors set in motion by internal programs.
Let us return to prey killing by cats. The cat’s brain must have a built-
The following experiment suggests the existence of such innate, internalized norms. Bryan Kolb and Arthur Nonneman allowed a litter of 6-
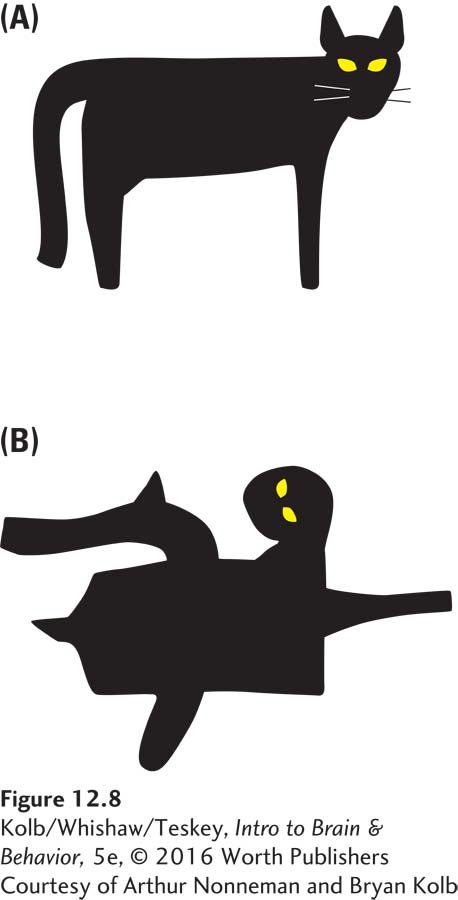
The kittens responded with raised fur, arched backs, and bared teeth, all signs of being threatened by the image of the adult. Some even hissed at the model. These kittens had no experience with any adult cat except their mother, and there was no reason to believe that she had ever shown them this behavior. Some sort of template of this posture must be prewired in the kitten brain. Seeing the model that matched this preexisting template automatically triggered a threat response. This innate trigger is an IRM.
The IRM concept also applies to humans. In one study, Tiffany Field and her colleagues (1982) had an adult display to young infants various exaggerated facial expressions, such as happiness, sadness, and surprise. As Figure 12-9 shows, the babies responded with very much the same expressions the adults displayed. These newborns were too young to be imitating the adult faces intentionally. Rather, babies must innately match these facial expressions to internal templates, in turn triggering some prewired program to reproduce the expressions in their own faces. Such an IRM would have adaptive value if these facial expressions serve as important social signals for humans.
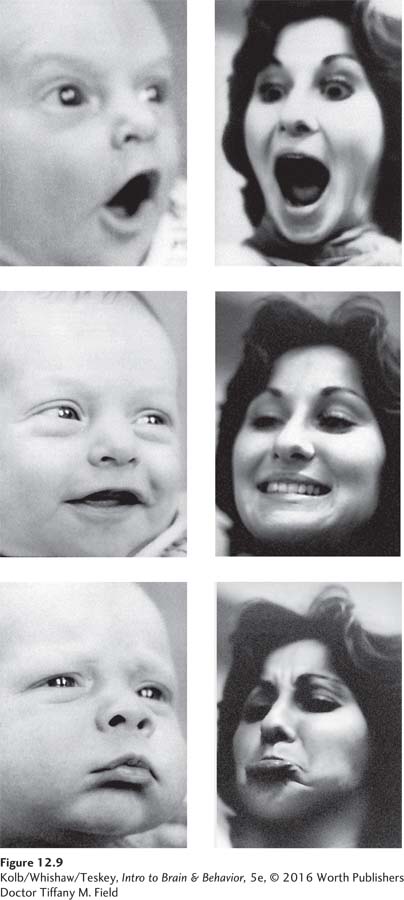
From M. Field, R. Woodson, R. Greenberg, D. Cohen, “Discrimination and imitation of facial expression by neonates,” Science 8 October 1982: Vol. 218 no. 4568 pp. 179–
Evidence for a prewired motor program related to facial expressions also comes from study of congenitally blind children, who spontaneously produce the very same facial expressions that sighted people do, even though they have never seen them in others. IRMs are prewired into the brain, but experience can modify them. Our cat Hunter’s stalking skills were not inherited fully developed at birth but rather matured functionally as she grew older. The same is true of many human IRMs, such as those for responding to sexually arousing stimuli.
Different cultures may emphasize different stimuli as arousing. Even within a single culture, what different people find sexually stimulating varies. Nonetheless, some human attributes are universally sexually arousing. For most human males, an example is the hip-
The IRM concept can be related to the Darwinian view of nervous system evolution. Natural selection favors behaviors that prove adaptive for an organism, and these behaviors are passed on to future generations. Because behavior patterns are produced by the activity of neurons in the brain, the natural selection of specific behaviors is really the selection of particular brain circuits.
Animals that survive long enough to reproduce and have healthy offspring are more likely to pass on their brain circuit genes than are animals with traits that make them less likely to survive and successfully reproduce. Thus, feral cats adept at stalking prey or responding fiercely to threats are more likely to survive and produce many offspring, passing on their adaptive brain circuits and behaviors to their young. In this way, the behaviors become widespread in the species over time.
The Darwinian view seems straightforward when we consider how cats evolved brain circuits for stalking prey or responding to threats. It is less obvious when applied to complex human behaviors, however. Why, for instance, have humans evolved the behavior of killing other humans? At first glance, it seems counterproductive to the survival of the human species. Why has it endured? For an answer, we turn to evolutionary psychology, the field that applies principles of natural selection to explanations of human behavior.
Evolutionary psychologists assume that any behavior, including homicide, occurs because natural selection has favored the neural circuits that produce it. When two men fight a duel, one commonsense explanation might be that they are fighting over grievances. But evolutionary psychologists would instead ask, why is a behavior pattern that risks people’s lives sustained in a population? Their answer: fights are about social status.
Men who fought and won duels passed on their genes to future generations. Through time, therefore, the traits associated with successful dueling—
Suppose that natural selection favored sexually jealous males who, to guarantee that they had fathered all offspring born of their mates, intimidated their rivals and bullied their mates. Male jealousy would be a prevalent motive for interpersonal violence, including homicide. Homicide itself does not help a man produce more children. But men who are apt to commit homicide are more likely to engage in other behaviors (bullying and intimidation) that improve their social status and therefore their reproductive fitness. Homicide therefore is related to adaptive traits that have been selected through millennia.
Section 15-5 posits a genetic explanation for the evolution of sex-
David Buss (2014) has examined patterns of mate selection across thousands of participants from 37 cultures, seeking to identify factors beyond culture that influence mate selection. His conclusions after nearly 30 years of study are that women around the world value dependability, stability, education, and intelligence in a long-
Older men and younger women are most likely to exhibit the mutually desired set of traits, which leads to a universal tendency for age differences between mates. Although the idea is controversial, Buss argues that these preferences are a product of natural selection in a Stone Age environment, when women and men would have faced different daily problems and thus would have developed separate adaptations related to mating.
Section 1-2 reviews Darwin’s theory, materialism, and contemporary perspectives on brain and behavior.
Evolutionary theory cannot account for all human behavior, perhaps not even homicide or mate selection. By casting an evolutionary perspective on the neurological bases of behavior, though, evolutionary psychologists can generate intriguing hypotheses about how natural selection might have shaped the brain and behavior.
Environmental Influences on Behavior
Many psychologists have emphasized learning as a cause of behavior. No one would question that we modify our behavior as we learn, but the noted behaviorist B. F. Skinner (1938) went much further. He believed that behaviors are selected by environmental factors.
Skinner’s argument is simple. Certain events function as rewards, or reinforcers. When a reinforcing event follows a particular response, similar responses are more likely to occur. Skinner argued that reinforcement can be manipulated to encourage the display of complex behaviors.

The power of experience to shape behavior by pairing stimuli and rewards is typified by one of Skinner’s experiments. A pigeon is placed in a box that has a small disc on one wall (the stimulus). If the pigeon pecks at the disc (the response), a food tray opens, and the pigeon can feed (the reinforcement or reward). The pigeon quickly learns the association between the stimulus and the response, especially if the disc has a small spot on it. It pecks at the spot and within minutes it has mastered the response needed to receive a reward.
Now the response requirement can be made more complex. The pigeon might be required to turn 360 degrees before pecking the disc to gain the reward. The pigeon can learn this response too. Other contingencies might then be added, making the response requirements even more complex. For instance, the pigeon might be trained to turn in a clockwise circle if the disc is green, to turn in a counterclockwise circle if the disc is red, and to scratch at the floor if the disc is yellow.
If you suddenly came across this complex behavior in a pigeon, you would probably be astounded. But if you understood the experience that had shaped the bird’s behavior, you would understand its cause. The rewards offered to the pigeon altered its behavior: its responses were controlled by the color of the disc on the wall.
Skinner extended behavioral analysis to include actions of all sorts—
Skinner also argued against the commonly held view that much of human behavior is under our own control. From Skinner’s perspective, free will is an illusion, because behavior is controlled not by the organism but rather by the environment, through experience. But what is the experience actually doing? Increasing evidence suggests that epigenetic changes regulate changes in memory circuits. Skinner was not studying the brain directly, but it is becoming clear that epigenetics supports his perspective. We learn many complex behaviors through changes in memory-
Epigenetic mechanisms mediate synaptic plasticity, especially in learning and memory. See Section 14-4.
The environment does not always change the brain. A case in point can be seen again in pigeons. A pigeon in a Skinner box can quickly learn to peck a disc to receive a bit of food, but it cannot learn to peck a disc to escape from a mild electric shock to its feet. Why not? Although the same simple pecking behavior is being rewarded, apparently the pigeon’s brain is not prewired for this second kind of association. The bird is prepared genetically to make the first association, for food, but not prepared for the second. This makes adaptive sense: typically, it flies away from noxious situations.
John Garcia and R. A. Koelling (1966) were the first psychologists to demonstrate the specific nature of this range of behavior–
So Garcia proposed an alternative to deter coyotes from killing lambs—
Many humans have similarly acquired food aversions because a certain food’s taste—
Section 14-4 has more on how our brains are wired to link unrelated stimuli.
One of us ate his first Caesar salad the night before coming down with a stomach flu. A year later, he was offered another Caesar salad and, to his amazement, felt ill just at the smell of it. Even though he knew that the salad had not caused his earlier illness, he nonetheless had formed an association between the novel flavor and the illness. This strong and rapid associative learning makes adaptive sense. Having a brain that is prepared to make a connection between a novel taste and subsequent illness helps an animal avoid poisonous foods and so aids in its survival. A curious aspect of taste aversion learning is that we are unaware of having formed the association until we encounter the taste and/or smell again.
The fact that the nervous system is often prewired to make certain associations but not to make others has led to the concept of preparedness in learning theories. Preparedness can help account for some complex behaviors. For example, if two rats are paired in a small box and exposed to a mild electric shock, they will immediately fight with one another, even though neither was responsible for the shock. Apparently, the rat brain is predisposed to associate injury with nearby objects or other animals. The extent to which we might extend this idea to explain such human behaviors as bigotry and racism is an interesting topic to ponder.
Inferring Purpose in Behavior: To Know a Fly
We may tend to assume that behavior is intentional. A wonderful little book, To Know a Fly, by Vincent Dethier (1962), disabuses this assumption by illustrating the problems in inferring purpose from an organism’s actions.
When a fly lands on a kitchen table, it wanders about, occasionally stomping its feet. Eventually, it finds a bit of food and sticks its proboscis (an extension like an elephant’s trunk) into the food and eats. The fly may then walk to a nearby place and begin to groom by rubbing its legs together quickly. Finally, it spends a long period motionless.
If you observed a fly engaged in these behaviors, it might appear to have been searching for food because it was hungry. When it found food, you might assume that it gorged itself until it was satisfied, then it cleaned up and rested. In short, the fly’s behavior might seem to you to be motivated—
Dethier studied flies for years to understand what a fly is actually doing when it engages in these behaviors. His findings have little to do with purpose or intention. When a fly wanders about a table, it is not deliberately searching. Rather, it is tasting what it walks on.
As Figure 12-10 shows, a fly’s taste receptors are on its feet. Tasting is automatic when a fly walks. An adult fly’s nervous system has a built-
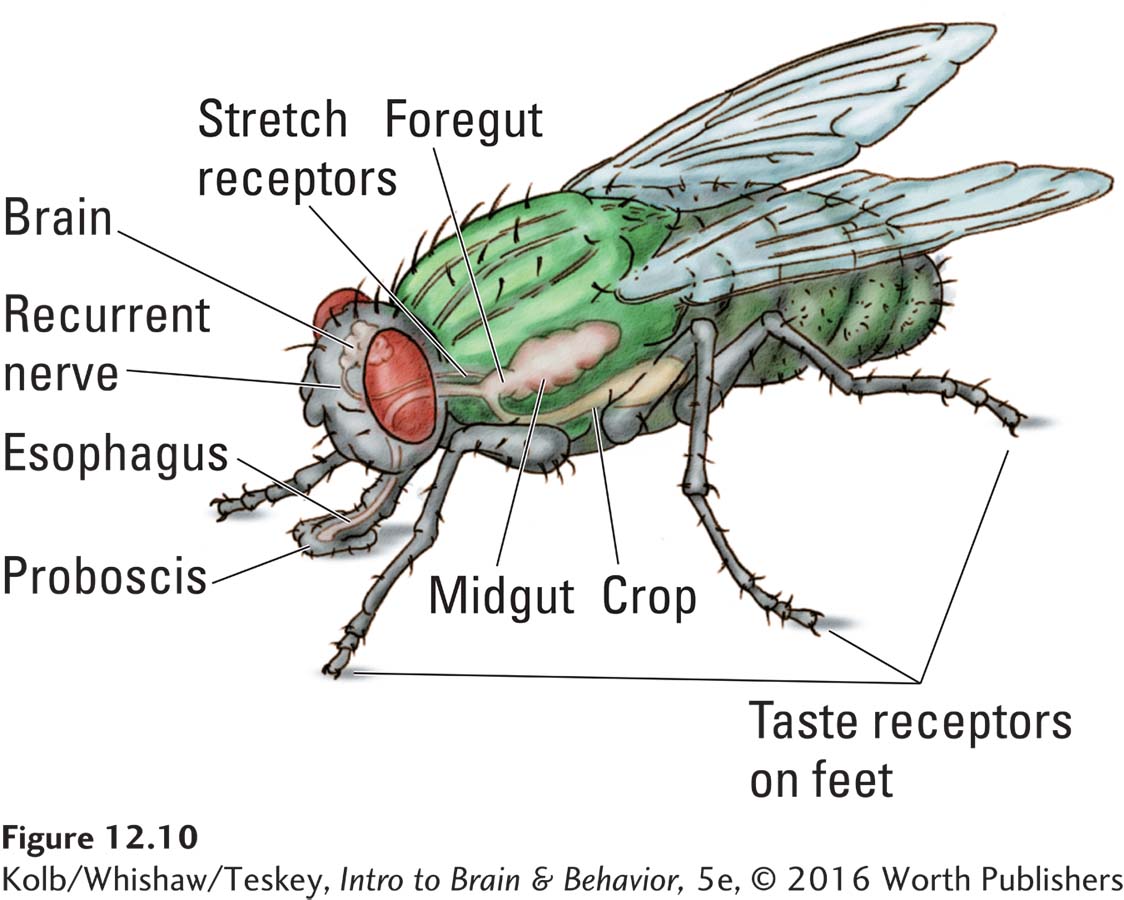
Why does a fly stop eating? A logical possibility is that its blood sugar level rises to some threshold. If this were correct, injecting glucose into the circulatory system of a fly would prevent the fly from eating. But that does not happen. Blood glucose level has no effect on a fly’s feeding. Furthermore, injecting food into the animal’s stomach or intestine has no effect either. So what is left?
Flies have a nerve (the recurrent nerve) that extends from the neck to the brain and carries information about whether any food is present in the esophagus. If the recurrent nerve is cut, the fly is chronically hungry and never stops eating. Such flies become so full and fat that their feet no longer reach the ground, and they become so heavy that they cannot fly.
Even though a fly appears to act with a purpose in mind, a series of very simple mechanisms actually control its behavior—
12-3 REVIEW
Evolution, Environment, and Behavior
Before you continue, check your understanding.
Question 1
B. F. Skinner argued that behaviors could be shaped by ______________ in the environment.
Question 2
John Garcia used the phenomenon of _______________ to discourage coyotes from killing lambs.
Question 3
The brain of a species is prewired to produce _____________ to specific sensory stimuli selected by evolution to prompt certain associations between events.
Question 4
When a fly wanders around on a table, it is not exploring so much as ____________.
Question 5
Explain briefly how the concept of preparedness accounts for puzzling human behaviors.
Answers appear in the Self Test section of the book.