14-1 Connecting Learning and Memory
The memory trace is a psychological construct, an abstract mental process inferred only from behavior (see Section 15-1). Others include learning, language, emotion, motivation, and thinking.
Learning is a relatively permanent change in an organism’s behavior as a result of experience. Memory is the ability to recall or recognize previous experience. Memory thus implies a mental representation of a previous experience, sometimes termed a memory trace. Neuroscientists presume that this hypothetical memory trace corresponds to a physical change in the brain, most likely involving synapses.
At the macro level, we infer what we know about learning and memory formation from behavioral changes, not by observing the brain directly. Studying learning and memory therefore requires behavioral measures that evaluate how these changes come about. We begin here by reviewing how learning and memory researchers study animals in the laboratory. The results suggest, in a general way, how the brain organizes its learning and memory systems.
Studying Learning and Memory in the Laboratory
A challenge for psychologists studying memory in laboratory animals (or people) is to get subjects to reveal what they remember. Because laboratory animals do not speak, investigators must devise ways for a subject to show its knowledge. Different species “talk” to us in different ways, so the test choice must match each species’ capabilities.
Figure 7-4 samples swimming pool tests for rats, Experiment 15-1, monkeys’ perceptual threshold, and Focus 15-4, the effects of brain injuries on humans’ cognitive performance.
Mazes or swimming pools are typically used to study rats because rats live in tunnels and near water. Monkey studies take advantage of their sharp vision and avid curiosity by requiring them to look under objects for food or watch television monitors. With birds, natural behaviors such as singing are used. And for humans, investigators tend to use paper-
Psychologists have devised hundreds of tests over the past century, and the test results reveal many types of learning and memory. Each appears to have its own neural circuitry. Two classic traditions for training animals to talk to investigators emerged a century ago. These diverse approaches are based on the work of Edward Thorndike (1898) in the United States and on experiments conducted by Ivan Pavlov in Russia.
Pavlovian Conditioning
Early in the twentieth century, Ivan Pavlov, a Russian physiologist, discovered that when a food reward accompanies some stimulus, such as a tone, dogs learn to associate the stimulus with the food. Then whenever they hear the tone, they salivate even though no food is present. This type of learning has many names, including Pavlovian conditioning, respondent conditioning, and classical conditioning, and many studies document its characteristics.
A key feature of Pavlovian conditioning is that animals learn to associate two stimuli (the presentation of the food and the tone) and to communicate to us that they have learned it by giving the same response (salivation) to both stimuli. Pet owners know that to a cat or dog, the sound of a can being opened is a clear stimulus for food. Two forms of Pavlovian conditioning are common in experiments today: eyeblink conditioning and fear conditioning. Each is associated with neural circuits in discrete brain regions; thus both are especially useful.
In the cerebellum, the flocculus controls eye movements; see Figure 11-14.
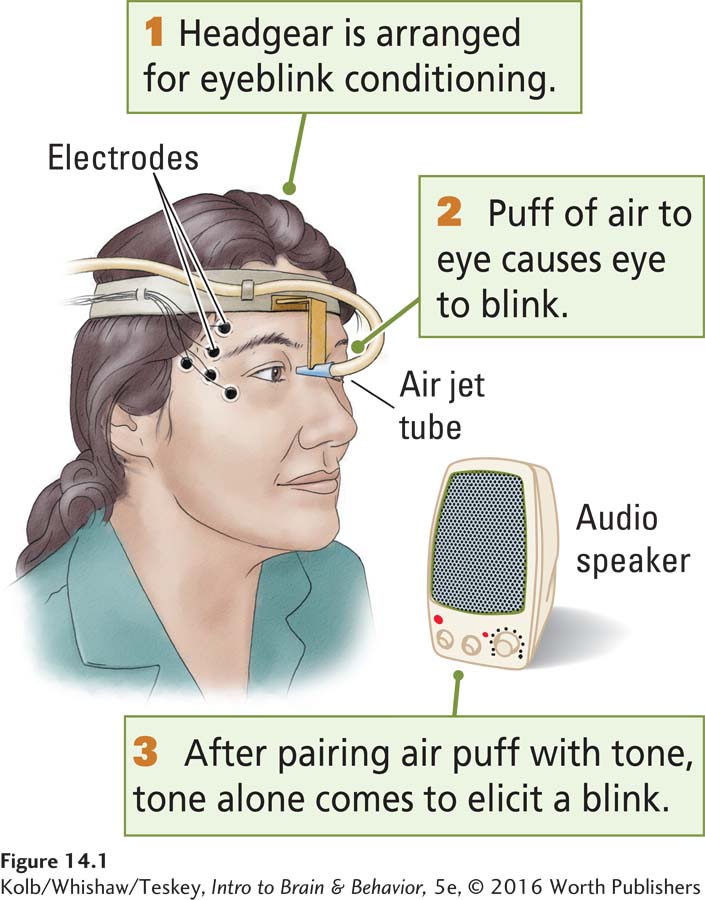
Eyeblink conditioning has been used to study Pavlovian learning in rabbits and people (Figure 14-1). In these studies, a tone (or some other stimulus) is associated with a painless puff of air to the participant’s eye. The tone is the conditioned stimulus (CS) that comes to elicit a blink produced initially by the air puff. The air puff is the unconditioned stimulus (UCS), because blinking is the normal reaction—
In fear conditioning, an unpleasant but harmless stimulus is used to elicit an emotional response: fear. A rat or other animal is placed in a box. A mild but unpleasant electric current can be passed through the grid floor. As shown in Experiment 14-1, a tone (the CS) is presented just before a brief, unexpected mild electric shock. When the tone is presented later without the shock, the animal acts afraid, becoming motionless and perhaps urinating in anticipation of the shock. A novel stimulus, say, a light, presented in the same environment has little effect. Thus, the animal communicates that it has learned the association between the tone and the shock.
EXPERIMENT
Question: Does an animal learn the association between emotional experience and environmental stimuli?
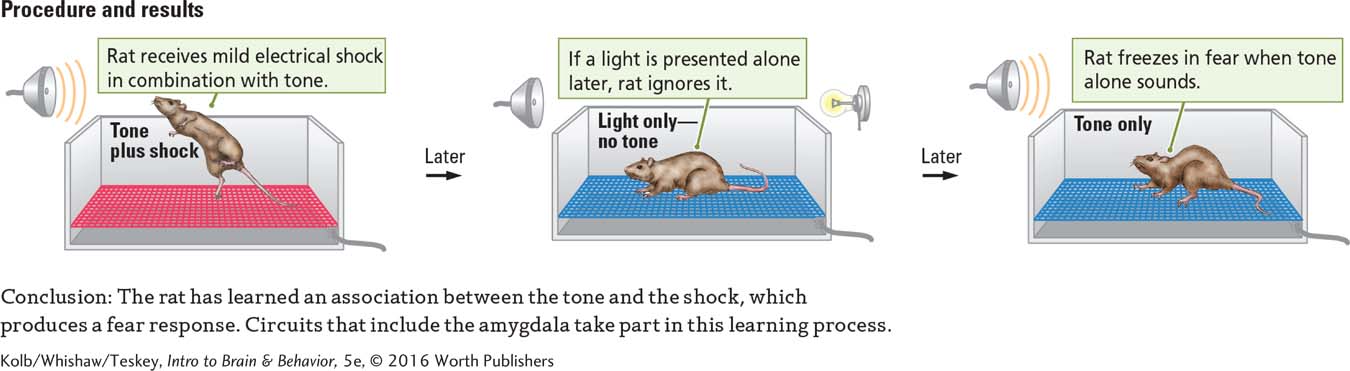
The shock approximates a spark of static electricity.
Because the CR is emotional, circuits of the amygdala rather than the cerebellum mediate fear conditioning. Although both eyeblink and fear conditioning are Pavlovian, different brain areas mediate the learning.
Operant Conditioning
In the United States, Edward Thorndike (1898) began a second tradition for studying learning and memory. Thorndike was interested in how animals solve problems. In one series of experiments, he placed cats in a box with a plate of fish outside it (Figure 14-2). The only way for a hungry cat to get to the fish was to figure out how to get out of the box.
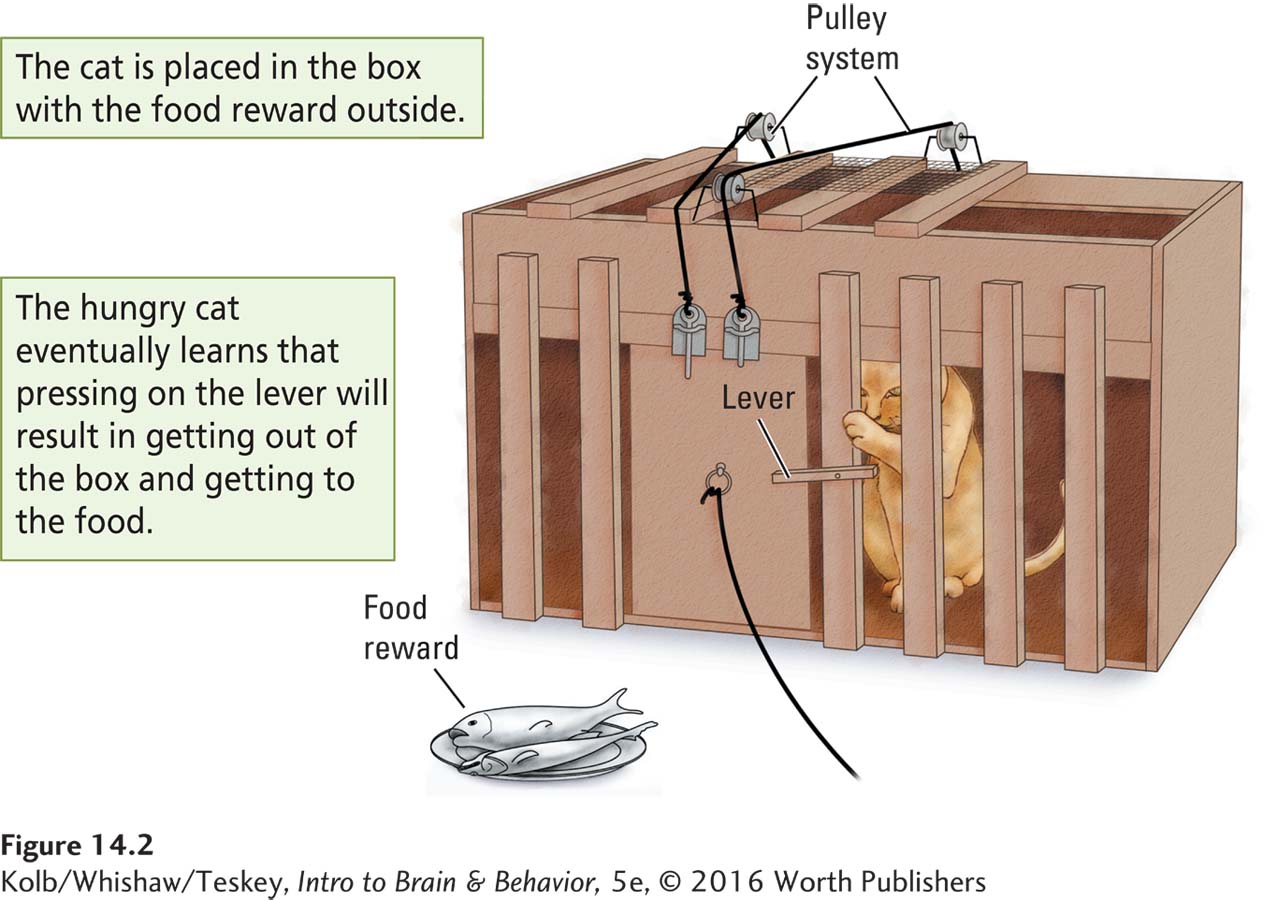
The solution was to press on a lever to activate a system of pulleys that opened the box door. The cat gradually learned that its actions had consequences: on the initial trial, the cat touched the releasing mechanism only by chance as it restlessly paced inside the box. The cat apparently learned that something it had done opened the door, and it tended to repeat its behaviors from just before the door opened. After a few trials, the cat took just seconds to get the door open to devour the fish.
Later studies by B. F. Skinner (e.g., 1938) used a similar strategy of reinforcement to train rats to press bars or pigeons to peck keys to obtain food. Just as Thorndike’s cats learned to escape his puzzle boxes, many animals learn to press the bar or peck the key simply if they are placed in the apparatus and allowed to discover the response that obtains the reward. This type of learning is operant conditioning, or instrumental conditioning, as Thorndike called it. The animal demonstrates that it has learned the association between its actions and the consequences by performing the task faster.
The variety of operant associations is staggering: we learn constantly to associate our behavior with its consequences. No surprise, then, that operant learning is not localized to any particular brain circuit. The circuits needed vary with the task requirements. For example, olfactory tasks involve olfactory-
Two Categories of Memory
Humans present a distinct challenge to researchers studying memory because so much of our learning is verbal. Psychologists have studied human memory since the mid-
In one kind of task, a group of participants reads a list of words, such as spring, winter, car, and boat. Another group reads a list consisting of trip, tumble, run, and sun. All the participants are then asked to define a series of words. One is fall.
The word fall has multiple meanings, including the season and a tumble. People who have just read the word list containing names of seasons are likely to give the meaning as autumn; those who have read the second list, containing action words, typically give the meaning as tumble. Some form of unconscious (and unintentional) learning takes place as the participants read the word lists.
This task measures implicit memory: participants demonstrate knowledge—
This implicit–

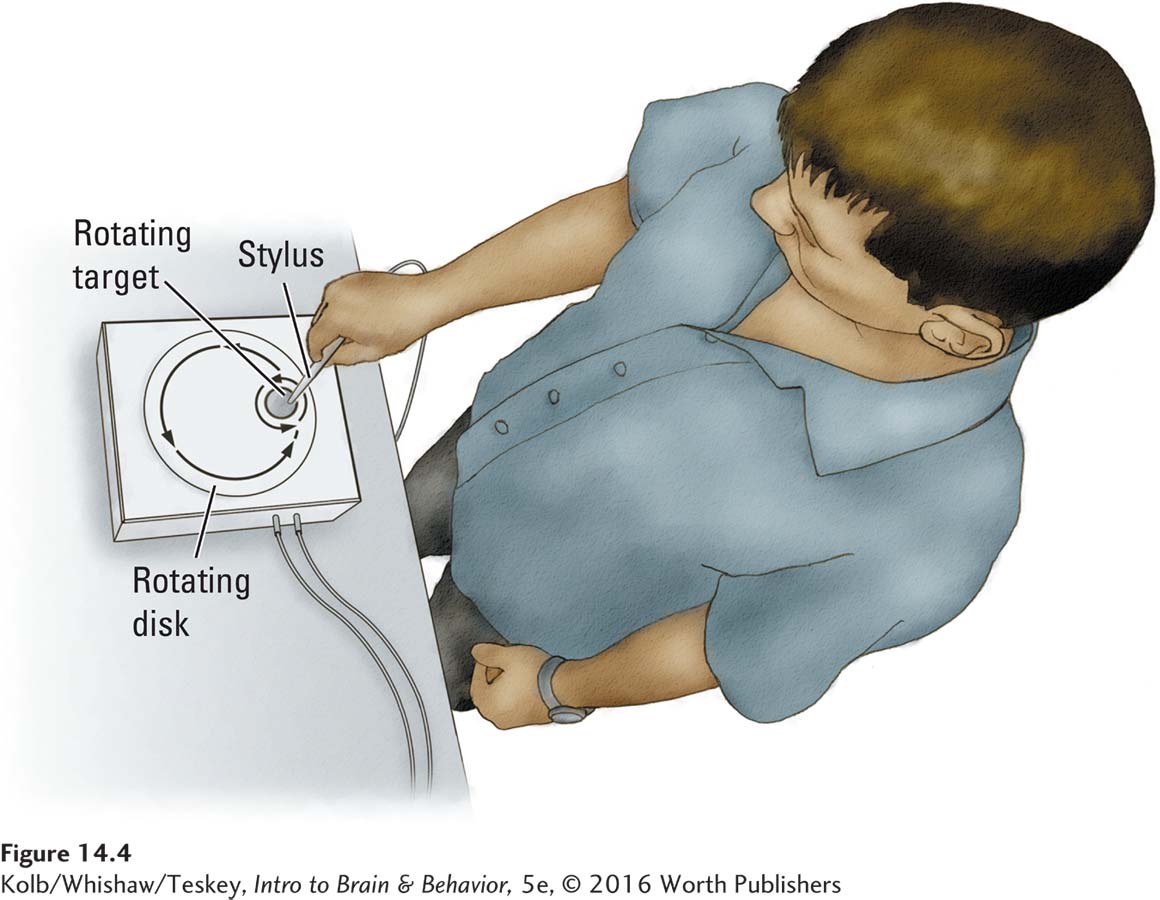
To measure implicit motor skills learning, a person learns a skill, such as the pursuit rotor task shown in Figure 14-4. A small metal disc moves in a circular pattern on a turntable that also is moving. The task is to hold a stylus on the small disc as it spins. This task is not as easy as it looks, especially when the turntable is moving quickly. Nonetheless, with an hour’s practice most people become reasonably proficient. Presented with the same task a week later, both controls and amnesics take less time to perform it. Here, too, the amnesics fail to recall performing the task before.
The distinction between tests of implicit and explicit memory is consistent and therefore must offer a key to how the brain stores information. Some theorists make subtle distinctions between the implicit–
Table 14-1 lists commonly used dichotomies, the general distinction being that one memory category requires recalling specific information, whereas the other refers to knowledge of which we are not consciously aware. We can include Pavlovian conditioning and Thorndike’s and Skinner’s operant learning in this analysis too: all are forms of implicit learning.
Nonspeaking animals can display explicit memory. One of us owned a cat that loved to play with a little ball. One day, as the cat watched, the ball was temporarily put on a high shelf to keep it away from an inquisitive toddler. For weeks afterward, the cat sat and stared at the shelf where the ball had been placed, even though the ball was not visible—
Animals also display explicit memory when they learn psychological tasks. Rats can be trained to find highly palatable food in a new location in a large compound each day. The task is to go to the most recent location. This piece of information is explicit and demonstrably can be forgotten.
Suppose a rat trained to find food in a particular location in a small arena is given several trials with the food at a new location and then retested an hour, a day, 3 days, or a week later. The rat has no difficulty with an hour’s delay or perhaps even a day’s. Some rats are flawless at 3 days, but most have forgotten the location by the time a week has elapsed. Instead, they wander around looking for the food. This behavior illustrates their implicit memory of the learning set, the rules of the game—
What Makes Explicit and Implicit Memory Different?
One reason explicit and implicit memories differ is that the set of neural structures that houses each is different. Another reason they differ is that the brain processes explicit and implicit information differently.
Encoding Memories
Implicit information is encoded in much the same way it is perceived: it can be described as data-
Explicit memory, in contrast, depends on conceptually driven, or top-
Because a person has a relatively passive role in encoding implicit memory, he or she has difficulty recalling the memory spontaneously but recalls it more easily with priming by the original stimulus or some feature of it. Because a person plays an active role in processing information explicitly, internal cues used in processing can also be used to initiate spontaneous recall.
Findings from studies of eyewitness testimony demonstrate the active nature of explicit memory recall—
Later questioning indicates that the memory of how fast the moving car was going is biased by the instruction: participants looking at “smashing” cars estimate faster speeds than those estimated by participants looking at “bumping” cars. The instruction actually causes the information to be processed differently. In both cases, participants were certain their memories were accurate.
Other experiments show that implicit memory also is fallible. For example, participants are read the following list of words: sweet, chocolate, shoe, table, candy, horse, car, cake, coffee, wall, book, cookie, hat. After a few minutes’ delay, they hear another list of words that includes some from the first list and some that are new. Participants are asked to identify which words were present on the first list and to indicate how certain they are of the identification.
One of the words on the second list is sugar. Most participants indicate not just that sugar was on the first list but that they are certain it was. Although other sweet things were, sugar was not. This demonstration is intriguing, because it shows how easily we can form false memories and defend their veracity with certainty.
Processing Memories
Although we can distinguish memories generally as implicit or explicit, the brain does not process all implicit or all explicit memories in the same way. Memories can be divided according to categories that differ from those listed in Table 14-1. For example, we can make a distinction between memories for different types of sensory information.
Different neural areas process visual and auditory information, so it is reasonable to assume that auditory memories are stored in brain regions different from the regions that store visual memories. We can also make a distinction between information stored in so-
The frontal lobes are central in short-
Storing Memories
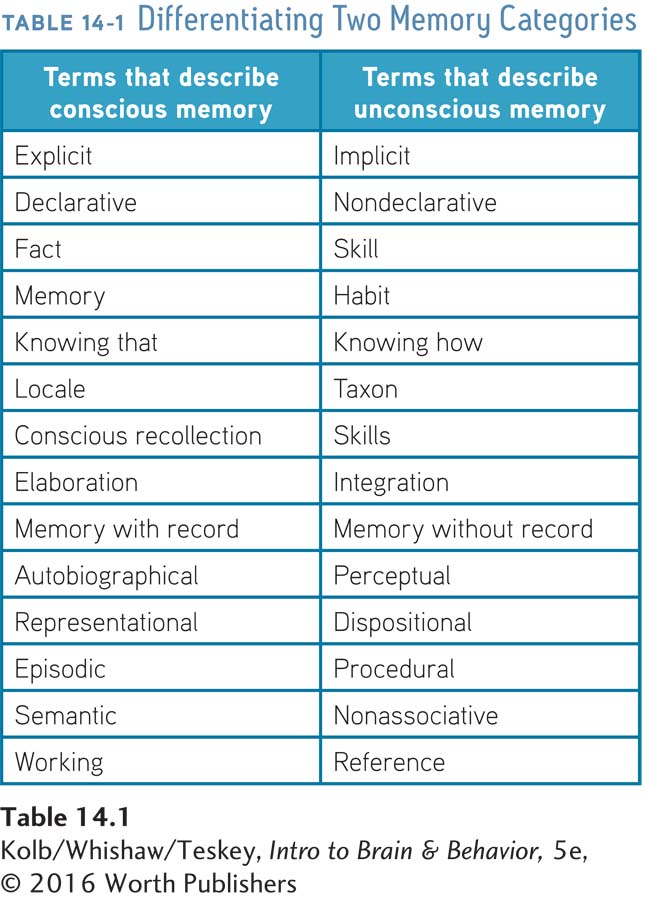
Understanding that every part of the brain can learn influences how we view the neural circuits that mediate memory. We could expect areas that process information to also house the memory of that information. Areas that process visual information, for example, probably house visual memory. Since the temporal lobe has specialized regions for processing color, shape, and other visual characteristics, we can predict that the memory for various visual attributes of objects is stored separately.
A series of PET studies by Alex Martin and colleagues (1995) at the U.S. National Institute of Mental Health confirmed this prediction. In one study, participants were shown black-
Just such dissociation was demonstrated. Figure 14-5 shows that color recall activates a region in the ventral temporal lobe, just anterior to the area controlling color perception, whereas recall of action words activates a region in the middle temporal gyrus, just anterior to the area controlling motion perception. This distribution of neural activation shows not only that object memory is at least partly located in the temporal lobes but also that it is found in regions associated with the original perception of the objects.
What Is Special about Personal Memories?
One aspect of memory unique to each of us is our personal, or autobiographical, memory. This episodic memory includes not only a record of events (episodes) that took place but also a record of our presence and role in the events. Our personal experiences form the basis of who we are and the rules by which we live. That is, we have memories not only for events but also for their context at a particular time in a particular place. We thus gain a concept of time and a sense of our personal role in a changing world.
Imagine what would happen if we lost our personal memories. We would still recall events but we would be unable to see our role in them. People with frontal lobe injuries sometimes exhibit such symptoms, as illustrated in a case described by Endel Tulving (2002).
K. C. suffered a serious traumatic brain injury in a motorcycle accident that produced multiple cortical and subcortical lesions. Remarkably, K. C.’s cognitive abilities were intact and indistinguishable from those of most typical healthy adults. He played chess and the organ, and his short-
What K. C. could not do was to recall any personally experienced events. This episodic amnesia covered his entire life, from birth. He knew facts about himself but had no memory for events that included him personally. For example, K. C. could not describe an event that took place in school that specifically included him, while at the same time recalling going to school and the knowledge he had gained there.
Findings from neuroimaging studies of people with episodic amnesia suggest that they consistently have frontal lobe injuries (Lepage et al., 2001), but exactly why these lesions produce episodic amnesia remains unclear. Nonetheless, Tulving made the interesting proposal that episodic memory is a marvel of nature: it transforms the brain into a kind of time machine that allows us to dwell on the past and make plans for the future. He goes further, suggesting that this ability may be unique to humans and is presumably due to some novel evolutionary development of the frontal lobe.
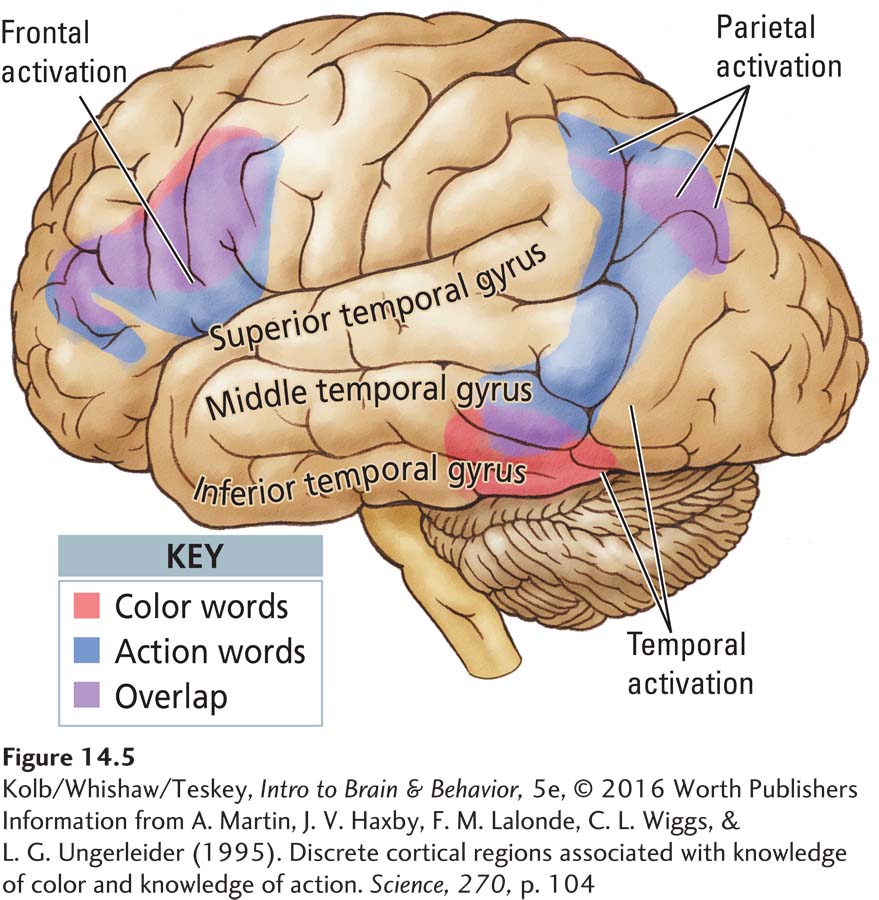
Not all people with episodic amnesia have brain injury, however. Many case reports describe patients with massive memory disturbances resulting from some “psychiatric” or “psychogenic” disorder. Such cases have been fodder for numerous movie plots. Hans Markowitsch (2003) noted that the amnesia reported in some of these cases is remarkably similar to episodic amnesia seen in neurological patients. Neuroimaging of patients with psychogenic amnesia shows a massive reduction in brain activity in frontal regions, a reduction remarkably similar to that seen in neurological patients with episodic amnesia (Figure 14-6). Therefore, we can assume that patients with psychogenic amnesias have a dysfunction of frontal brain activity that blocks the retrieval of autobiographical memory.
Just as some people exhibit poor autobiographical memory, a rare group displays highly superior autobiographical memory (HSAM) (LePort et al., 2012). These people display virtually complete recall for events in their lives, usually beginning around age 10, and can often describe any episode, including the day of the week that it occurred and the date. They can even recall the weather that day, as well as social and public events. Brain imaging of those who display HSAM shows increased gray matter in the temporal and parietal lobes and increased size in the fiber projection between the temporal and frontal lobes.
Lawrence Pathihis and colleagues (2013) wondered if HSAM individuals might also be immune to memory distortion, such as the false memories described earlier. The investigators found that HSAM individuals are as likely to develop false memories as are other participants. Whatever the source of their extraordinary autobiographical memory, it does not prevent the sort of memory distortions the rest of us experience. Possibly that is because the research paradigms employed are not strictly autobiographical.
14-1 REVIEW
Connecting Learning and Memory
Before you continue, check your understanding.
Question 1
An organism learns that some stimulus is paired with a reward. This is ____________ conditioning.
Question 2
After learning that consequences follow its behavior, an organism modifies its behavior. This is ____________ conditioning.
Question 3
Information that is unconsciously learned forms ____________ memory, whereas specific factual information forms ____________ memory.
Question 4
____________ memory is autobiographical and unique to each person.
Question 5
Where is memory stored in the brain?
Answers appear in the Self Test section of the book.