The Resting Membrane Potential in Animal Cells Depends Largely on the Outward Flow of K+ Ions Through Open K+ Channels
The plasma membranes of animal cells contain many open K+ channels but few open Na+,Cl−, or Ca2+ channels. As a result, the major ionic movement across the plasma membrane is the movement of K+ from the inside outward, powered by the K+ concentration gradient. This movement leaves an excess of negative charge on the cytosolic face of the plasma membrane and creates an excess of positive charge on the exoplasmic face, as in the experimental system shown in Figure 11-18c. This outward flow of K+ ions is the major determinant of the inside-negative membrane potential. The channels through which the K+ ions flow, called resting K+ channels, alternate, like all channels, between an open and a closed state (see Figure 11-2), but since their opening and closing is not affected by the membrane potential or by small signaling molecules, these channels are referred to as nongated. In contrast, the various gated channels in neurons and other excitable cells (see Chapter 22) open only in response to specific ligands or to changes in membrane potential.
Quantitatively, the usual resting membrane potential of –60 to –70 mV is close to the potassium equilibrium potential, calculated from the Nernst equation and the K+ concentrations in cells and surrounding media (see Table 11-2). Usually the resting membrane potential is slightly lower (less negative) than that calculated from the Nernst equation because of the presence of a few open Na+ channels. These channels allow the net inward flow of Na+ ions, making the cytosolic face of the plasma membrane more positive—that is, less negative—than predicted by the Nernst equation for K+. The K+ concentration gradient that drives the flow of ions through resting K+ channels is generated by the Na+/K+ ATPase described previously (see Figures 11-3 and 11-13). In the absence of this pump, or when it is inhibited, the K+ concentration gradient cannot be maintained, the membrane potential falls to zero, and the cell eventually dies.
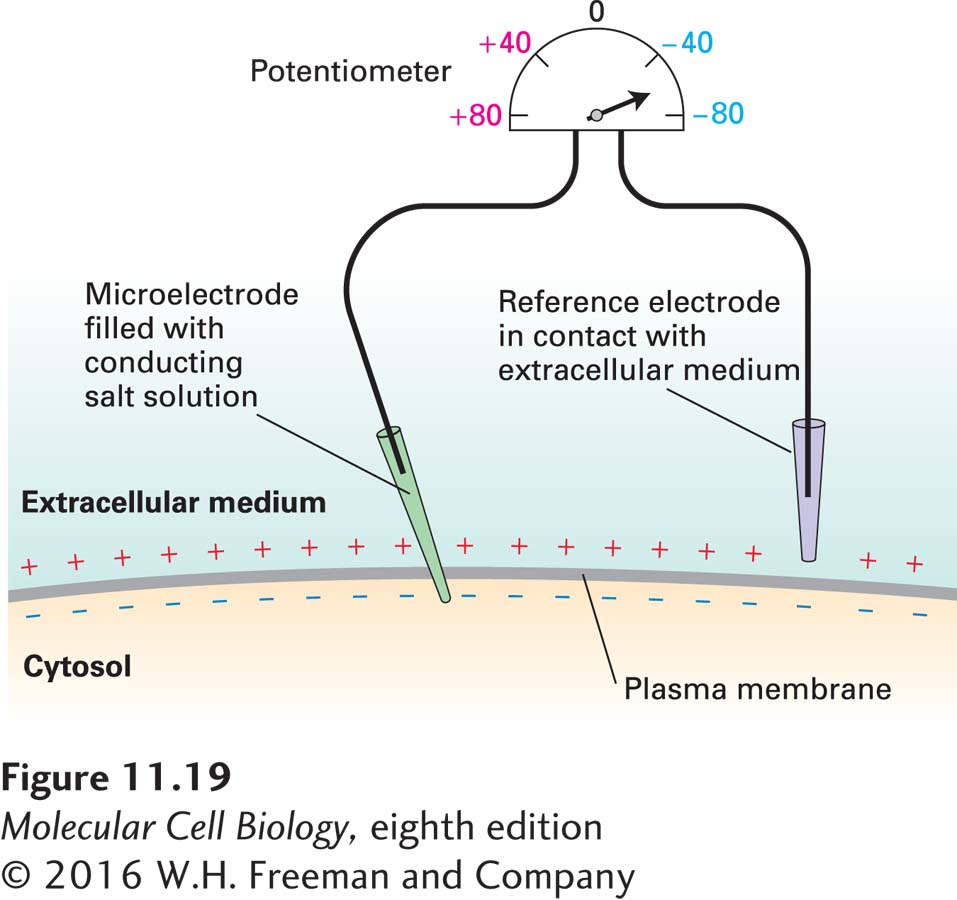
EXPERIMENTAL FIGURE 11-19The electric potential across the plasma membrane of a live cell can be measured. A microelectrode, constructed by filling a glass tube of extremely small diameter with a conducting fluid such as a KCl solution, is inserted into a cell in such a way that the plasma membrane seals itself around the tip of the electrode. A reference electrode is placed in the extracellular medium. A potentiometer connecting the two electrodes registers the potential—in this case, –60 mV, with the cytosolic face negative with respect to the exoplasmic face of the membrane. A potential difference is registered only when the microelectrode is inserted into the cell; no potential is registered if the microelectrode is in the extracellular fluid.
Although resting K+ channels play the dominant role in generating the electric potential across the plasma membranes of animal cells, this is not the case in bacterial, plant, and fungal cells. The inside-negative membrane potential in plant and fungal cells is generated by transport of positively charged protons (H+) out of the cell by ATP-powered proton pumps, a process similar to what occurs in lysosomal membranes lacking Cl− channels (see Figure 11-14a): each H+ pumped out of the cell leaves behind a Cl− ion, generating an inside-negative electric potential across the membrane. In aerobic bacterial cells, an inside-negative potential is generated by outward pumping of protons during electron transport, a process similar to proton pumping in mitochondrial inner membranes that will be discussed in detail in Chapter 12 (see Figure 12-22).
The electric potential across the plasma membrane of a cell can be measured with a microelectrode inserted into the cell and a reference electrode placed in the extracellular fluid. The two electrodes are connected to a potentiometer capable of measuring small potential differences (Figure 11-19). The potential across the plasma membrane of most animal cells generally does not vary with time. In contrast, neurons and muscle cells—the principal types of electrically active cells—undergo controlled changes in their membrane potential, as we discuss in Chapter 22.