Mitochondria Are Influenced by Direct Contacts with the Endoplasmic Reticulum
Mitochondrial dynamics, and indeed, many mitochondrial functions, are influenced by direct contacts between mitochondria and the endoplasmic reticulum (ER). The portions of the ER that form special contact regions with the mitochondria, called mitochondria-associated membranes, or MAMs, can be visualized using electron microscopy and fluorescence microscopy (Figure 12-13). Their lipid and protein composition differs somewhat from that of the rest of the ER. In yeast, a protein complex called ERMES (ER-mitochondria encounter structure) has been proposed to mediate the reversible tethering of MAMs to mitochondria. The ERMES complex is not present in mammals; the proteins that mediate the tethering of MAMs to mitochondria in complex multicellular organisms are as yet unknown. Tethering proteins hold the MAM and the outer mitochondrial membrane about 10–30 nm apart.
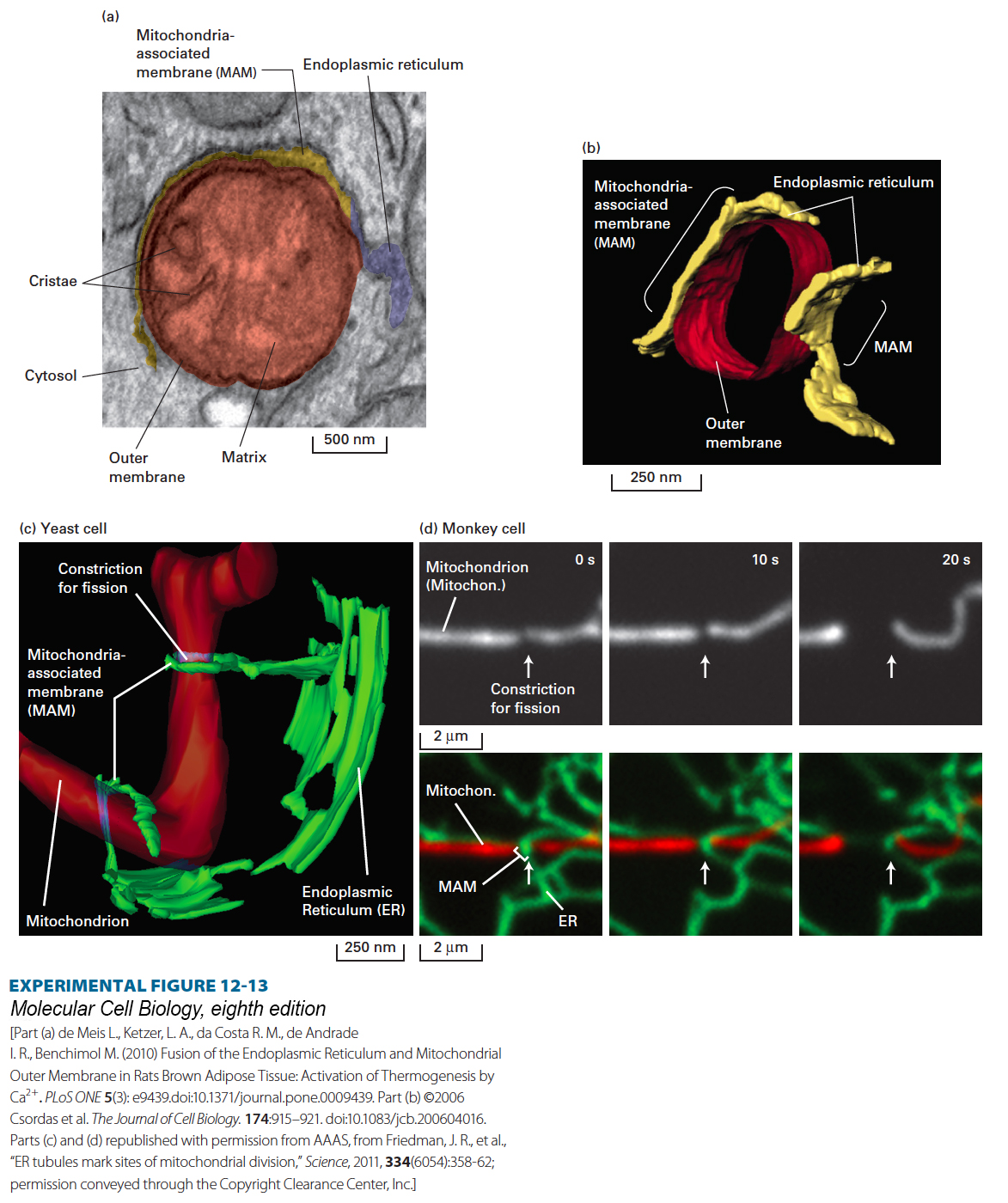
EXPERIMENTAL FIGURE 12-13Specialized regions of the endoplasmic reticulum called mitochondria-associated membranes (MAMs) directly contact mitochondria and influence mitochondrial shape, function, and sites of fission. (a) Transmission electron microscopic (EM) image of a section through rat brown adipose (fat) tissue. The lumen of the endoplasmic reticulum (ER) is false colored to show a MAM (yellow) and the non-MAM, bulk ER (blue). The MAM is closely apposed to the outer mitochondrial membrane. (b) Three-dimensional model of a segment of a mitochondrion (red, only outer membrane shown) and the adjacent MAM (yellow) determined from a line of cultured avian lymphoma cells using EM tomography (assembly of a three-dimensional image from consecutive individual sections). (c) A three-dimensional model of a mitochondrion (red) and adjacent MAMs (green) from a yeast cell using EM tomography. The two MAM domains are derived from ER tubules that in some cases can wrap around the mitochondrion, in the top case forming a clamp-like structure that appears to constrict the mitochondrion in preparation for fission. (d) Live cell fluorescence microscopic images of a Cos-7 monkey cell, showing a mitochondrion (white in the top panels, same mitochondrion in red in the bottom panels) and MAM (green in bottom panels), taken from a single field of view at 10-second intervals. The arrow points to the site of constriction and fission on the mitochondrion and to the MAM at the constriction/fission site. The MAM directs constriction and subsequent DRP1-mediated fission at this site. To visualize the mitochondria and ER, the Cos-7 cells were transfected with cDNA vectors encoding two fluorescent proteins that specifically accumulate in either the mitochondrion (red fluorescence) or the ER (green fluorescence).
MAMs contribute significantly to many cellular processes (see Table 12-1), including mitochondrial fission. MAM-mitochondrial contacts can initiate mitochondrial constriction and help recruit DPR1, which completes membrane fission (see Figure 12-12c). In yeast, MAM tubules have been seen to loop completely around mitochondria, forming a clamp that constricts the mitochondrion (Figure 12-13c). In mammalian cells, the MAMs contact the mitochondria at fission sites, but they have not been shown to loop fully around the mitochondria (Figure 12-13d).
MAMs also play an integral role in intracellular calcium and energy metabolism. Variations in the concentrations of calcium ions in intracellular compartments—cytosolic calcium ([Ca2+]c), mitochondrial calcium ([Ca2+]m), and calcium in the ER ([Ca2+]er)—are employed to control a wide variety of activities within cells, a process called calcium signaling (see Chapter 15). Calcium is also important for extracellular processes, such as the activity of some blood-clotting proteins. Intramitochondrial calcium ions play an important role in controlling mitochondrial function, and MAMs mediate this control by delivering calcium from the ER to mitochondria. For example, an increase in [Ca2+]m in the matrix can increase mitochondrial production of ATP. Increased [Ca2+]m directly increases the activities of three mitochondrial enzymes that produce NADH from NAD+: pyruvate dehydrogenase (see Figure 12-5) and α-ketoglutarate and isocitrate dehydrogenases (see Figure 12-16 below). As we shall see later in this chapter, NADH provides high-energy electrons for ATP synthesis. Thus continuous low-level release of Ca2+ from MAMs into mitochondria is necessary for ATP synthesis when cells are in a basal, or resting, state. Increased delivery of Ca2+ via MAMs can occur when cells require more ATP—for example, when muscle cells are stimulated to contract. Strikingly, calcium signaling is used both to induce muscle contraction (see Chapter 17) and coordinately to increase mitochondrial ATP synthesis to provide the energy to fuel that contraction. When [Ca2+]m is elevated, mitophagy can be induced. Indeed, mitochondrial calcium overload can activate regulated cell death pathways. Thus the control of [Ca2+]m can literally control the life and death of cells.