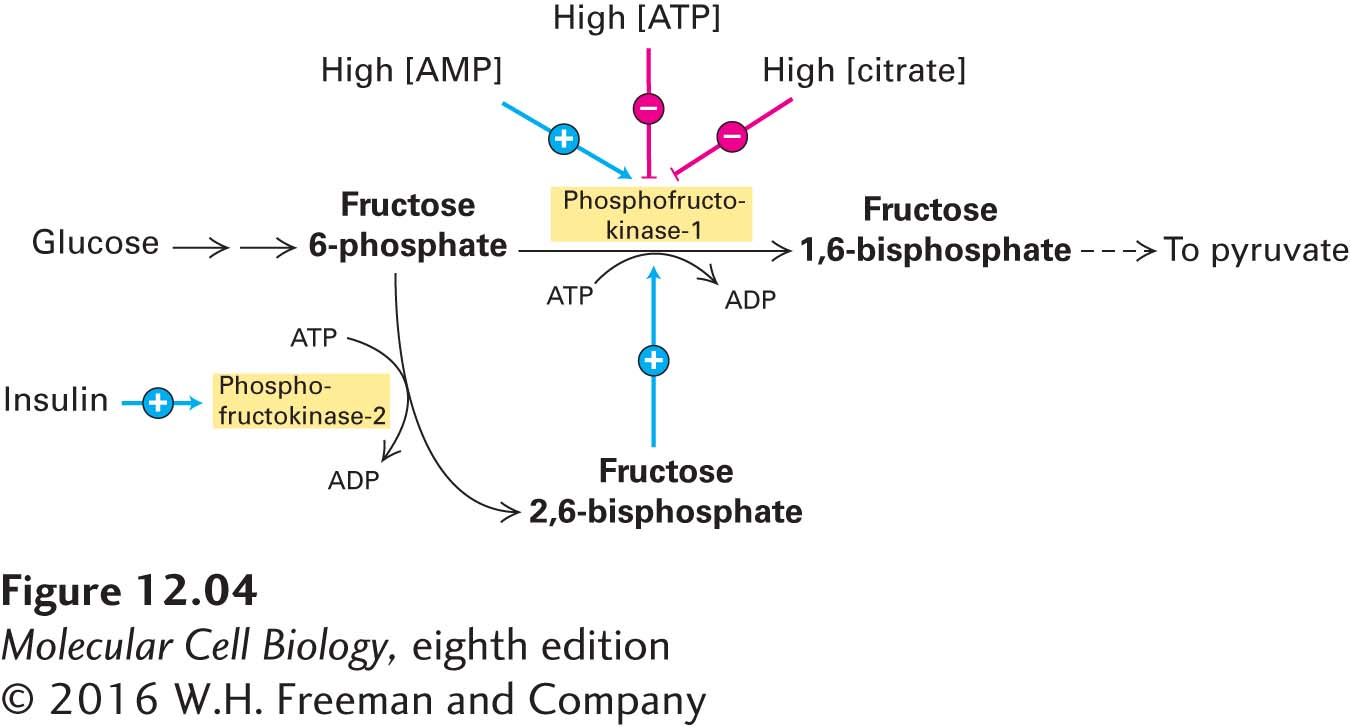
The Rate of Glycolysis Is Adjusted to Meet the Cell’s Need for ATP
To maintain appropriate levels of ATP, cells must control the rate of glucose catabolism. The operation of the glycolytic pathway (stage I), as well as the citric acid cycle (stage II), is continuously regulated, primarily by allosteric mechanisms (see Chapter 3 for general principles of allosteric control). Three allosteric enzymes involved in glycolysis play key roles in regulating the entire glycolytic pathway. Hexokinase (see Figure 12-3, step 1) is inhibited by its reaction product, glucose 6-phosphate. Pyruvate kinase (step 10) is inhibited by ATP, so glycolysis slows down if too much ATP is present. The third enzyme, phosphofructokinase-1 (step 3), is the principal rate-limiting enzyme of the glycolytic pathway. In a manner that is emblematic of its critical role in regulating the rate of glycolysis, this enzyme is allosterically controlled by several molecules (Figure 12-4).
FIGURE 12-4Allosteric regulation of glucose metabolism. The key regulatory enzyme in glycolysis, phosphofructokinase-1, is allosterically activated by AMP and fructose 2,6-bisphosphate, which are elevated when the cell’s energy stores are low. The enzyme is inhibited by ATP and citrate, both of which are elevated when the cell is actively oxidizing glucose to CO2 (i.e., when energy stores are high). Later we will see that citrate is generated during stage II of glucose oxidation. Phosphofructokinase-2 is a bifunctional enzyme: its kinase activity forms fructose 2,6-bisphosphate from fructose 6-phosphate, and its phosphatase activity catalyzes the reverse reaction. Insulin, which is released by the pancreas when blood glucose levels are high, promotes phosphofructokinase-2 kinase activity and thus stimulates glycolysis. At low blood glucose, glucagon is released by the pancreas and promotes phosphofructokinase-2 phosphatase activity in the liver, indirectly slowing down glycolysis.
Page 517
For example, phosphofructokinase-1 is allosterically inhibited by ATP and allosterically activated by adenosine monophosphate (AMP). As a result, the rate of glycolysis is very sensitive to the cell’s energy charge, a measure of the fraction of total adenosine phosphates that have “high-energy” phosphoanhydride bonds, which is equal to [(ATP) + 0.5 (ADP)]/[(ATP) + (ADP) + (AMP)]. The allosteric inhibition of phosphofructokinase-1 by ATP may seem unusual because ATP is also a substrate of this enzyme. But the affinity of the substrate-binding site for ATP is much higher (has a lower Km) than that of the allosteric site. Thus at low concentrations, ATP binds to the catalytic site, but not to the inhibitory allosteric site, and enzymatic catalysis proceeds at near-maximal rates. At high concentrations, ATP also binds to the allosteric site, inducing a conformational change that reduces the affinity of the enzyme for its other substrate, fructose 6-phosphate, and thus reduces the rate of this reaction and the overall rate of glycolysis.
Another important allosteric activator of phosphofructokinase-1 is fructose 2,6-bisphosphate. This metabolite is formed from fructose 6-phosphate by an enzyme called phosphofructokinase-2. Fructose 6-phosphate accelerates the formation of fructose 2,6-bisphosphate, which in turn activates phosphofructokinase-1. This type of control is known as feed-forward activation, in which a high abundance of a metabolite (here, fructose 6-phosphate) accelerates its subsequent metabolism. Fructose 2,6-bisphosphate allosterically activates phosphofructokinase-1 in liver cells by decreasing the inhibitory effect of high ATP concentrations and by increasing the affinity of phosphofructokinase-1 for one of its substrates, fructose 6-phosphate.
The three glycolytic enzymes that are regulated by allostery catalyze reactions with large negative ΔG°′ values—reactions that are essentially irreversible under ordinary conditions. These enzymes are therefore particularly suitable for regulating the entire glycolytic pathway. Additional control is exerted by glyceraldehyde 3-phosphate dehydrogenase, which catalyzes the reduction of NAD+ to NADH (see Figure 12-3, step 6). As we shall see, NADH is a high-energy electron carrier used subsequently during oxidative phosphorylation in mitochondria. If cytosolic NADH builds up owing to a slowdown in mitochondrial oxidation, step 6 becomes thermodynamically less favorable.
Page 518
Glucose metabolism is controlled differently in various mammalian tissues to meet the metabolic needs of the organism as a whole. During periods of carbohydrate starvation, for instance, it is necessary for the liver to release glucose into the bloodstream. To do this, the liver converts the polymer glycogen, a storage form of glucose (see Chapter 2), directly to glucose 6-phosphate (without involvement of hexokinase, step 1). Under these conditions, there is a reduction in fructose 2,6-bisphosphate levels and decreased phosphofructokinase-1 activity (see Figure 12-4). As a result, glucose 6-phosphate derived from glycogen is not metabolized to pyruvate; rather, it is converted to glucose by a phosphatase and released into the blood to nourish the brain and red blood cells, which depend primarily on glucose for their energy. In all cases, the activity of these regulated enzymes is controlled by the level of small-molecule metabolites, generally by allosteric interactions, or by hormone-mediated phosphorylation and dephosphorylation reactions. (Chapter 15 gives a more detailed discussion of hormonal control of glucose metabolism in liver and muscle.)