Chromosomes Are Captured and Oriented During Prometaphase
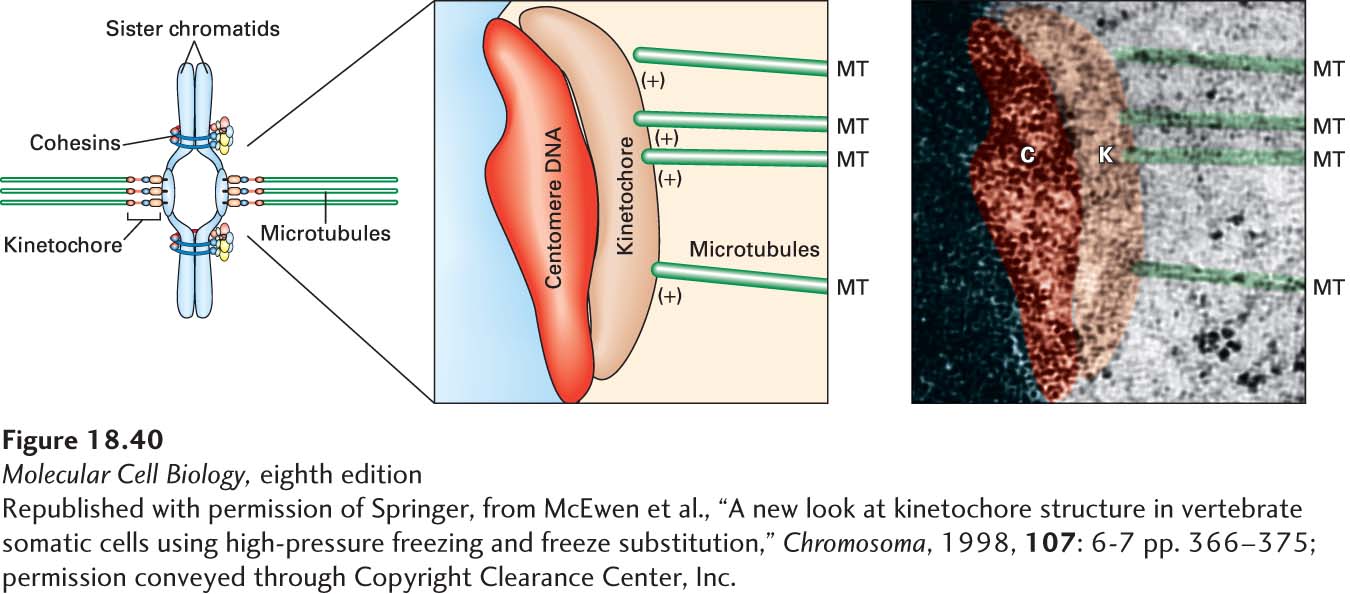
Kinetochores, the structures that mediate attachment between chromosomes and microtubules, assemble on each sister chromatid at a region called the centromere. The centromere is a constricted region of the condensed chromosome defined by a centromeric DNA sequence. Centromeric DNA can vary enormously in size; in budding yeast it is about 125 bp, whereas in humans it is on the order of 1 Mb (see Chapter 8). Kinetochores contain many protein complexes to facilitate the linkage between centromeric DNA and microtubules. In animal cells, the kinetochore consists of a centromeric DNA layer and inner and outer kinetochore layers, with the (+) ends of kinetochore microtubules terminating in the outer layer (Figure 18-40). Yeast kinetochores are attached by a single microtubule to their spindle pole, human kinetochores are attached by about 30 microtubules, and plant chromosomes by hundreds.
[Republished with permission of Springer, from McEwen et al., “A new look at kinetochore structure in vertebrate somatic cells using high-pressure freezing and freeze substitution,” Chromosoma, 1998, 107: 6-7 pp. 366-375; permission conveyed through Copyright Clearance Center, Inc.]
FIGURE 18-40The structure of a mammalian kinetochore. Diagram and electron micrograph of a mammalian kinetochore.
[Republished with permission of Springer, from McEwen et al., “A new look at kinetochore structure in vertebrate somatic cells using high-pressure freezing and freeze substitution,” Chromosoma, 1998, 107: 6-7 pp. 366-375; permission conveyed through Copyright Clearance Center, Inc.]
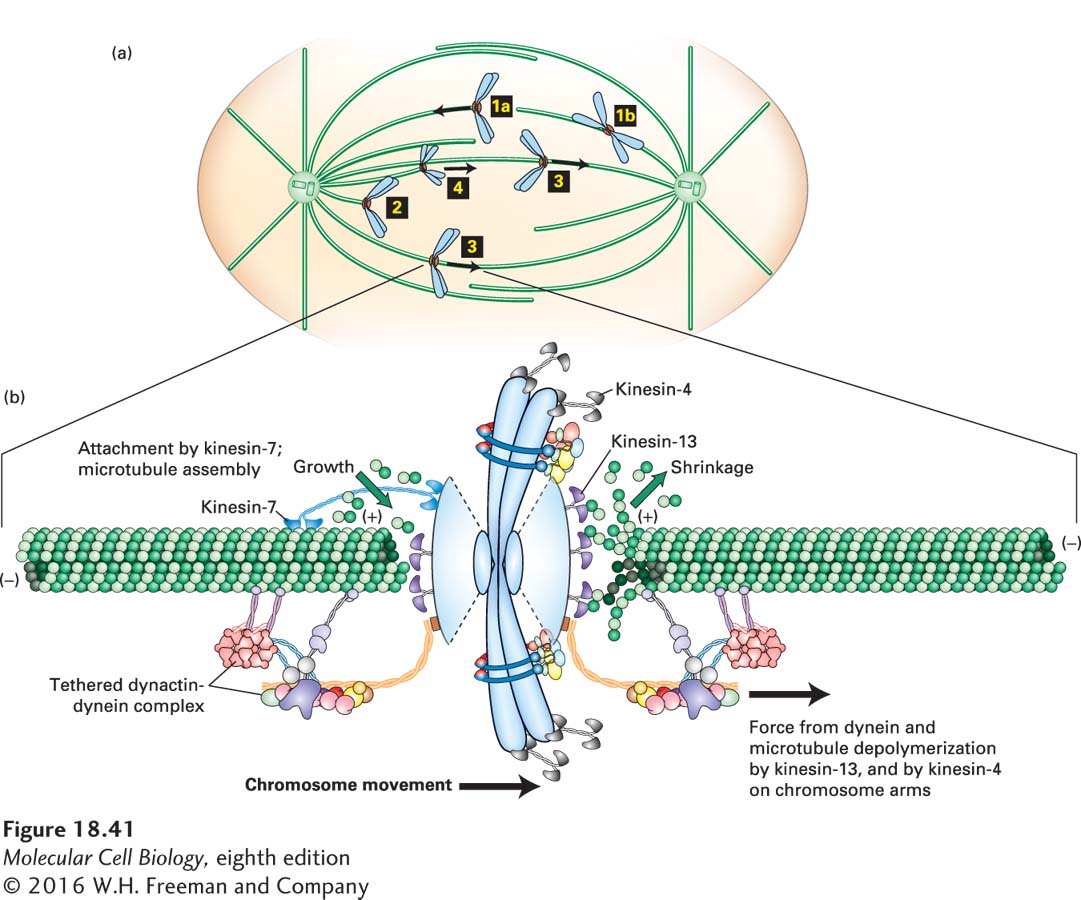
How does a kinetochore become attached to microtubules in prometaphase? Microtubules nucleated from the spindle poles are very dynamic, and when they contact a kinetochore, either laterally or at their ends, this contact can lead to chromosomal attachment (Figure 18-41a, steps 1a and 1b). Microtubules that “capture” kinetochores are selectively stabilized by a reduction in the rate of catastrophes, which increases the chance that the attachment will persist.
FIGURE 18-41Chromosome capture and congression in prometaphase. (a) In the first stage of prometaphase, chromosomes become attached either to the end of a microtubule (1a) or to the side of a microtubule (1b). The chromosome is then drawn toward the spindle pole by dynein-dynactin that becomes associated with one of the kinetochores of the chromosome and moves toward the (−) end of the microtubule (2). Eventually, a microtubule from the opposite pole finds and becomes attached to the free kinetochore, and the chromosome is now said to be bi-oriented (3). Once some chromosomes are bi-oriented, others, having established one kinetochore-pole interaction, use CENP-E/kinesin-7 on their free kinetochore to aid in orientation (4). The bi-oriented chromosomes then move to a central point between the spindle poles in a process known as chromosome congression. Note that during these steps, chromosome arms point away from the closest spindle pole: this is due to chromokinesin/kinesin-4 motors on the chromosome arms moving toward the (+) ends of the polar microtubules. In animal cells, many microtubules associate with each kinetochore. For ease of presentation, only single kinetochore microtubules are shown here. (b) Congression involves bidirectional oscillations of chromosomes, with one set of kinetochore microtubules shortening on one side of the chromosomes and the other set lengthening on the other. On the shortening side, a kinesin-13 protein stimulates microtubule disassembly and a dynein-dynactin complex moves the chromosome toward the pole. On the side with lengthening microtubules, kinesin-7 protein holds on to the growing microtubule. The kinetochore also contains many additional protein complexes not shown here. See Cleveland et al., 2003, Cell112:407–421.
Page 854
Recent studies have uncovered a mechanism involving Ran, a small GTPase, that enhances the chance that microtubules will encounter kinetochores. Recall from Chapter 13 that during interphase, the Ran GTPase cycle is involved in the transport of proteins into and out of the nucleus through nuclear pores (see Figure 13-37). During mitosis, when the nuclear membrane and pores have disassembled, a guanine nucleotide exchange factor for the Ran GTPase is bound to chromosomes, thereby generating a higher local concentration of Ran⋅GTP in the vicinity of the chromosomes. Because the enzyme that stimulates GTP hydrolysis on Ran—the Ran GAP—is evenly distributed in the cytosol, this generates a gradient of Ran⋅GTP centered on the chromosomes. Ran⋅GTP induces the association of cytosolic microtubule-stabilizing factors with the microtubule, resulting in enhanced microtubule growth, and in this way biases growth of microtubules nucleated from spindle poles toward chromosomes.
Once a kinetochore is attached laterally or terminally to a microtubule, dynein-dynactin associates with the kinetochore to move the duplicated chromosome down the microtubule toward the spindle pole. This movement eventually results in an end-on attachment of the microtubule to one kinetochore (Figure 18-41a, step 2). This movement helps orient the sister chromatid so that the unoccupied kinetochore on the opposite side is pointing toward the distal spindle pole. Eventually a microtubule from the distal pole will capture the free kinetochore; at this point the sister chromatid pair is said to be bi-oriented (Figure 18-41a, step 3). With the two kinetochores attached to opposite poles, the duplicated chromosome is now under tension, being pulled in both directions by the two sets of kinetochore microtubules. When one or a few chromosomes are bi-oriented, other chromosomes use these existing kinetochore microtubules to contribute to their orientation and movement to the spindle center. This orientation is mediated by kinesin-7 (also known as CENP-E) associated with the free kinetochore, which moves the chromosome to the (+) end of the kinetochore microtubule (Figure 18-41a, step 4).