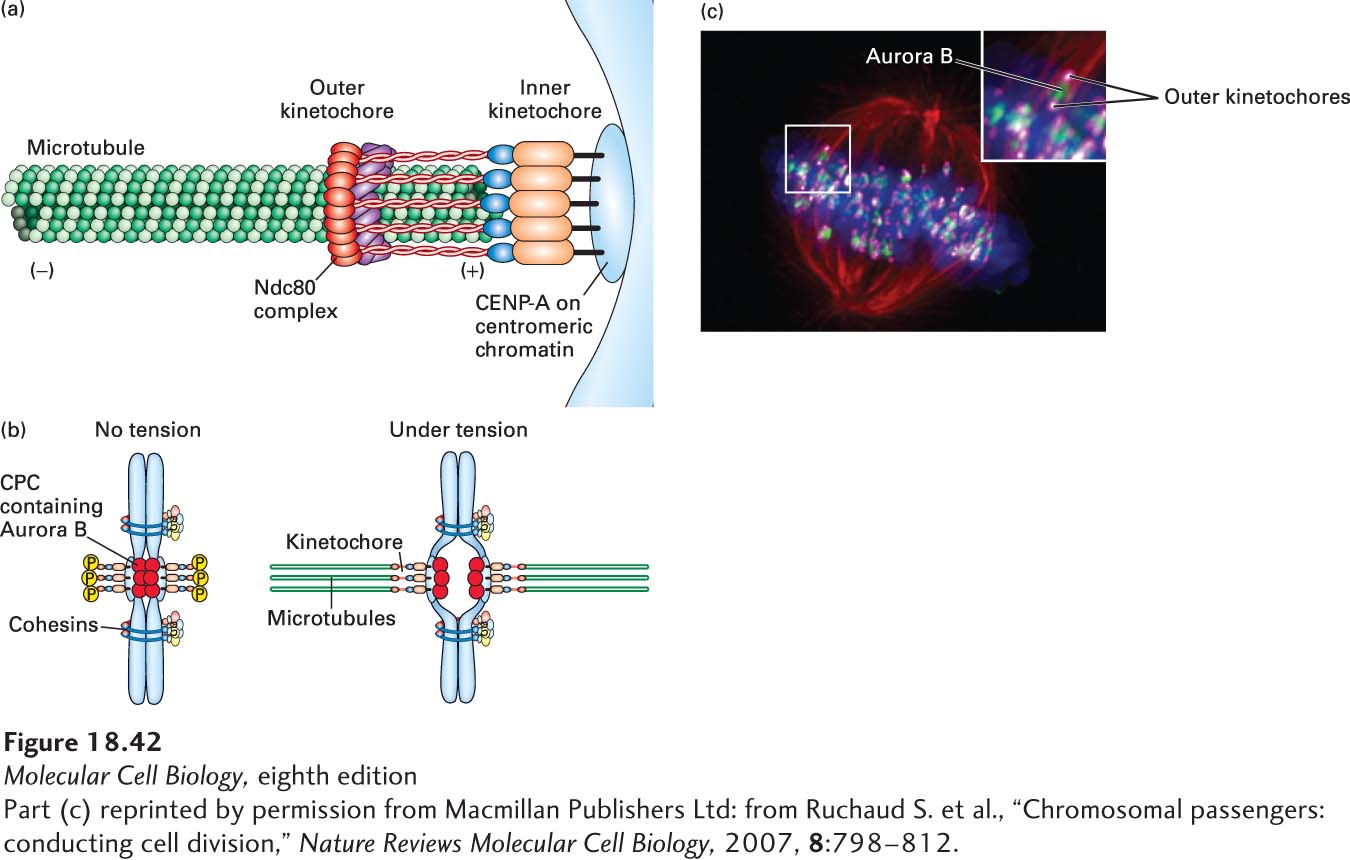
The Chromosomal Passenger Complex Regulates Microtubule Attachment at Kinetochores
We have noted that the segregation of chromosomes at mitosis must be very accurate, so it is crucial that all chromosomes are bi-oriented before anaphase begins. During the random kinetochore-to-microtubule attachment process, it is possible for mistakes to be made; for example, both kinetochores of a sister chromatid pair might attach to microtubules from the same spindle pole. If such attachments persisted during metaphase, it would result in one cell missing a chromosome and the other having an extra one, which would either be lethal or very detrimental. Cells have two important mechanisms to ensure that all chromosomes are correctly bi-oriented before anaphase begins.
The first mechanism ensures that the kinetochore-microtubule interactions are weak until bi-orientation occurs. When a chromosome is correctly bi-oriented, tension is produced across the chromosome, and this tension leads to the kinetochore-microtubule attachments becoming stabilized. To understand how this works, we need to look a bit more closely at the molecular components that link a kinetochore to a microtubule. As we discussed in Chapter 8, kinetochores assemble on regions of chromosomal DNA marked by a centromere-specific H3 histone variant called CENP-A. This variant marks the site for kinetochore assembly, which is a very complicated process. About half a dozen distinct stable protein complexes, consisting of more than 40 different proteins, have been shown to associate with this centromeric region in yeast. Essentially all these protein complexes are conserved in humans, which is not surprising given the fundamental importance of kinetochores. One of these, the so-called Ndc80 complex, is long and flexible, and many copies of it link the inner kinetochore with the (+) end of the microtubule in a sleevelike arrangement (Figure 18-42a). The function of Ndc80 and many of the associated factors at the kinetochore is regulated by the chromosomal passenger complex (CPC). This complex associates with the inner kinetochore early in mitosis, and among its components is a protein kinase called Aurora B. Once the CPC is associated with the kinetochore, Aurora B can phosphorylate several components in the near vicinity, including the Ndc80 complex, which loosens the attachment of Ndc80 to the microtubule. The phosphorylation of these components is not stable, however: another protein, the phosphatase PP1, which is associated with the outer kinetochore, can dephosphorylate them. Thus when the kinetochores on a pair of sister chromatids are not under tension, Ndc80 is continually phosphorylated by Aurora B and dephosphorylated by PP1. The result is a weak interaction between the kinetochore and microtubule. However, when bi-orientation occurs, the tension generated pulls on both kinetochores and extends the flexible Ndc80 complex to increase the spacing between the inner and outer kinetochores (Figure 18-42b, c). As a result of these movements, Ndc80 cannot be phosphorylated by Aurora B, and the dephosphorylated state of Ndc80 renders it more firmly attached to the microtubule. In this way, microtubule attachments to bi-oriented chromosomes are selectively stabilized.
[Part (c) reprinted by permission from Macmillan Publishers Ltd: from Ruchaud S. et al., “Chromosomal passengers: conducting cell division,” Nature Reviews Molecular Cell Biology, 2007, 8:798–812.]
FIGURE 18-42CPC regulation of microtubule-kinetochore attachment. The Ndc80 complex forms a critical and regulated attachment between the kinetochore and microtubule (+) end. (a) Diagram showing the sleevelike arrangement of the Ndc80 complex linking the inner kinetochore to the (+) end of a microtubule embedded in the outer kinetochore. See S. Santaguida and A. Musacchio, 2009, EMBO J.28:2511–2531. (b) Diagram of the relationship between the chromosomal passenger complex (CPC), which is associated with the inner kinetochore and contains the kinase Aurora B, and the outer kinetochore, where the phosphatase PP1 binds. Notice that when both kinetochores are under tension, the outer kinetochores move away from the CPC; as a result, Aurora B cannot phosphorylate components in the outer kinetochore, which include the microtubule-binding site of the Ndc80 complex. (c) Cell in metaphase stained for tubulin (red), DNA (blue), Aurora B kinase (green), and the outer kinetochore (magenta). Notice how the outer kinetochore is pulled away from Aurora B (inset).
[Part (c) reprinted by permission from Macmillan Publishers Ltd: from Ruchaud S. et al., “Chromosomal passengers: conducting cell division,” Nature Reviews Molecular Cell Biology, 2007, 8:798–812.]
Page 856
While the CPC is important for bi-orientation of each individual chromosome, it does not ensure that all chromosomes are bi-oriented before anaphase begins. The second mechanism to ensure correct chromosome segregation is the spindle assembly checkpoint pathway, a signaling pathway that stops the progression of the cell cycle into anaphase until tension is present at all the kinetochores. Even a single unattached, or inappropriately attached, kinetochore can activate the spindle assembly checkpoint pathway and pause the cell cycle until the error is corrected. This mechanism, discussed in detail in Chapter 19, guarantees that all the chromosomes are correctly bi-oriented before the cell proceeds into anaphase.