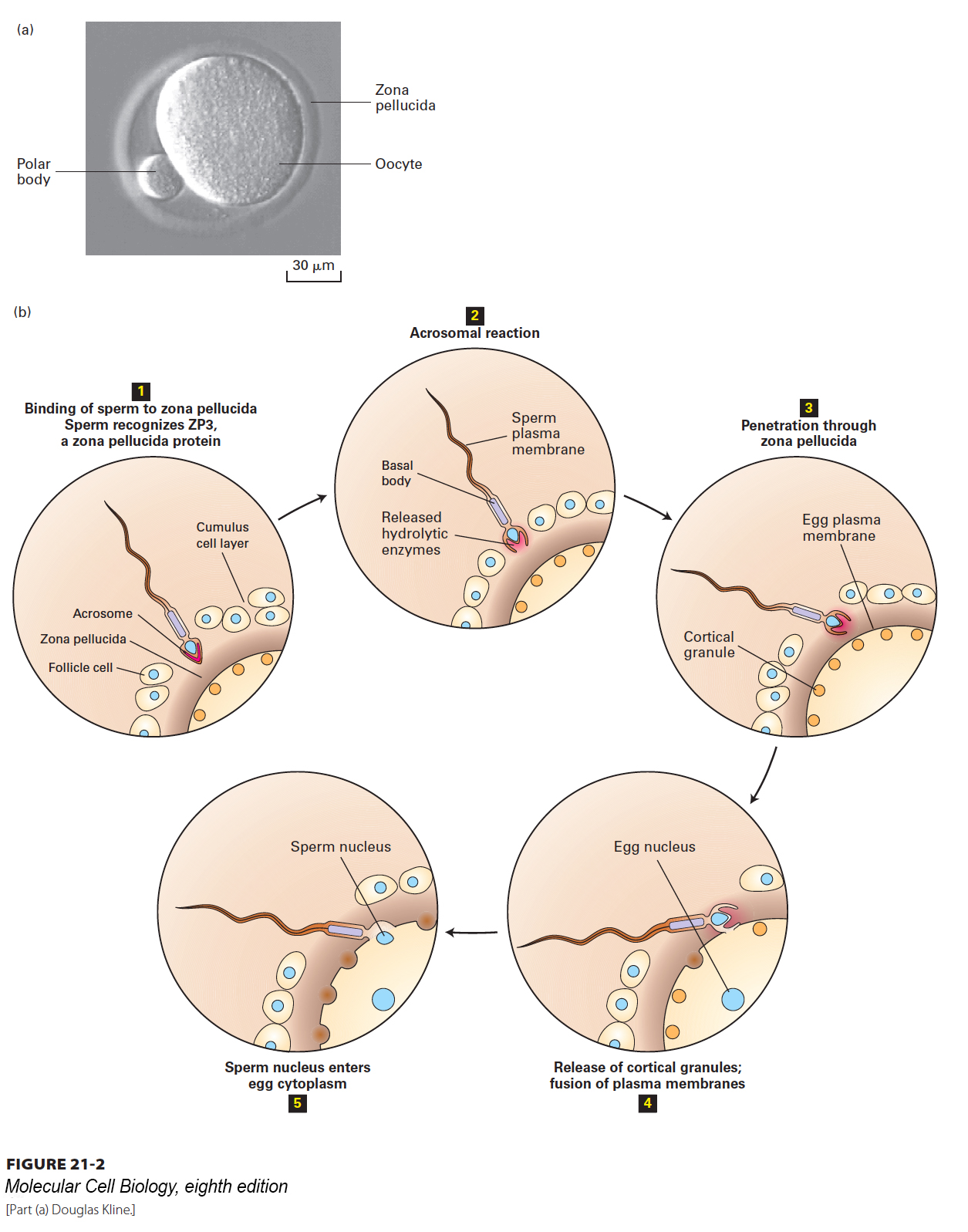
It is remarkable that a mammalian sperm is ever able to reach and penetrate an egg. For one thing, in humans, each sperm is competing with more than 100 million other sperm for a single oocyte. What’s more, the sperm must swim an incredible distance to reach the egg (if a sperm were the size of a person, the distance traveled would be equivalent to several miles!). And once there, the sperm must fight its way through multiple layers surrounding the egg that restrict sperm entry (Figure 21-2a). Sperm are streamlined for speed and swimming ability. The human sperm flagellum (see Chapter 18) contains about 9000 dynein motors that flex microtubules in the 50-µm axoneme. Nevertheless, only a few dozen sperm will reach the oocyte.
[Part (a) Douglas Kline.]
FIGURE 21-2Gamete fusion during fertilization. (a) Mammalian eggs, such as the mouse oocyte shown here, are surrounded by a ring of translucent material, the zona pellucida, which provides a binding matrix for sperm. The diameter of a mouse egg is ~70 µm, and the zona pellucida is ~6 µm thick. The polar body is a nonfunctional product of meiosis. (b) In the initial stage of fertilization (step 1), the sperm penetrates a layer of cumulus cells surrounding the egg to reach the zona pellucida. Interactions between GalT, a protein on the sperm surface, and ZP3, a glycoprotein in the zona pellucida, trigger the acrosomal reaction (step 2), which releases enzymes from the acrosome. Degradation of the zona pellucida by hydrolases and proteases released by the acrosomal reaction allows the sperm to begin entering the egg (step 3). Specific recognition proteins on the surfaces of egg and sperm facilitate fusion of their plasma membranes. Membrane fusion and the subsequent entry of the first sperm nucleus into the egg cytoplasm (steps 4 and 5) trigger the release of Ca2+ within the oocyte. Cortical granules (orange) respond to the Ca2+ surge by fusing with the oocyte membrane and releasing enzymes that act on the zona pellucida to prevent binding of additional sperm.
[Part (a) Douglas Kline.]
As shown in Figure 21-2, once it reaches the egg, a sperm must first penetrate a layer of cumulus cells that surround the oocyte and then the zona pellucida, a gelatinous extracellular matrix composed largely of three glycoproteins called ZP1, ZP2, and ZP3. The acrosome, found at the sperm’s leading tip, is a membrane-bounded compartment specialized for interaction with the oocyte. One side of the acrosomal membrane lies just under the plasma membrane at the sperm head; the opposite side of the acrosomal membrane is juxtaposed to the nuclear membrane. Inside the acrosome are soluble enzymes, including hydrolases and proteases. Once in proximity to the oocyte, the acrosome undergoes exocytosis, releasing its contents onto the surface of the oocyte (Figure 21-2b, step 2). The enzymes digest the multiple egg surface layers to begin the process of sperm entry. It’s a race, and the first sperm to succeed triggers a dramatic response by the oocyte that prevents polyspermy, the entry of other sperm that would bring in excess chromosomes.
Page 978
Page 979
After the first sperm succeeds in fusing with the oocyte membrane, a flux of calcium flows into the oocyte cytosol, spreading outward from the site of sperm entry. As in other regulated secretory pathways, one of the effects of the rise in calcium is to trigger fusion of vesicles located just under the plasma membrane of the egg, called cortical granules, with the plasma membrane, releasing their contents to the outside of the plasma membrane and forming a shielding fertilization membrane that blocks other sperm from entering. Finally the sperm nucleus enters the egg cytoplasm, and the egg and sperm nuclei soon fuse to create the diploid zygote nucleus.
Oocytes bring with them to the union a considerable dowry. They contain multiple mitochondria, with their mitochondrial DNA, whose inheritance is exclusively maternal; in mammals and many other species, no sperm mitochondrial DNA enters the oocyte (see Chapter 12). Female-specific mitochondrial DNA inheritance has been used to trace maternal heritage in human history; it has been used, for example, to follow early humans from their origins in Africa. The egg cytoplasm is also packed with maternal mRNA: transcripts of genes whose products are essential for the earliest stages of development. There is little or no transcription during oocyte meiosis and the first embryonic cleavages, so during this time the oocyte’s mRNA is crucial.