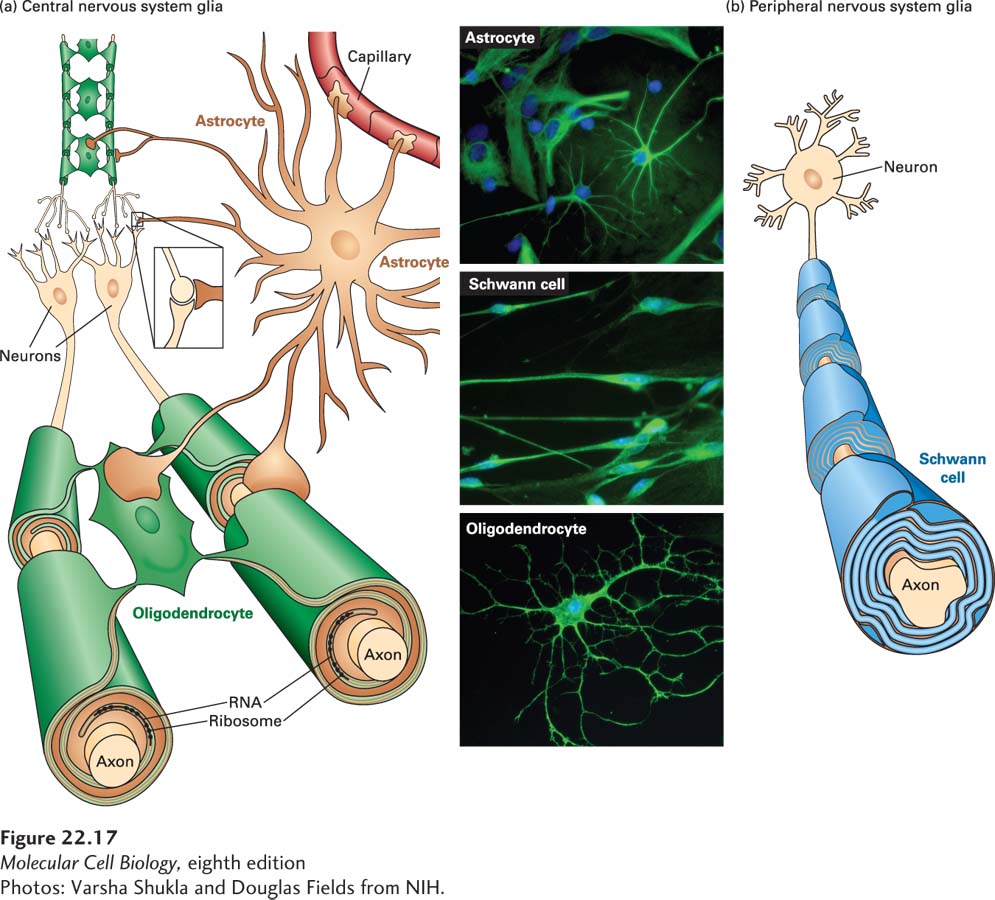
Figure 22-17 shows three main types of glial cells present in the nervous system, two of which produce myelin sheaths: oligodendrocytes make sheaths for the central nervous system (CNS), and Schwann cells make them for the peripheral nervous system (PNS). Astrocytes, also shown in the figure, facilitate synapse formation and communication between neurons, and are discussed in Sections 22.1 and 22.3. A fourth type of glia, microglia (not shown), constitutes a part of the CNS immune system. While microglia are not related by lineage to neurons or to other glia, they have recently been shown to play roles in an aspect of neural circuit formation called synaptic pruning, discussed in Section 22.3.
[Photos: Varsha Shukla and Douglas Fields from NIH.]
FIGURE 22-17Three types of glial cells. (a) A single oligodendrocyte in the central nervous system can myelinate segments of multiple axons. Astrocytes interact with neurons but do not form myelin. (b) Each Schwann cell insulates a section of a single peripheral nervous system axon. See B. Stevens, 2003, Curr. Biol.13:R469, and D. L. Sherman and P. Brophy, 2005, Nature Rev. Neurosci. 6:683–690.
[Photos: Varsha Shukla and Douglas Fields from NIH.]
Oligodendrocytes Oligodendrocytes form the spiral myelin sheath around axons of the central nervous system (Figure 22-17a). Each oligodendrocyte provides myelin sheaths to segments of multiple neurons. The major protein constituents are myelin basic protein (MBP) and proteolipid protein (PLP). MBP, a peripheral membrane protein found in both the central and peripheral nervous systems (Figure 22-18), has seven RNA splicing variants that encode different forms of the protein. It is synthesized by ribosomes located in the growing myelin sheath, an example of specific transport of mRNAs to a distal cell region (Chapter 10). MBP mRNA undergoes microtubule-dependent transport to distal oligodendrocyte processes, where its local translation contributes to the formation of the myelin sheath.
[Part (a) ISM/Phototake.]
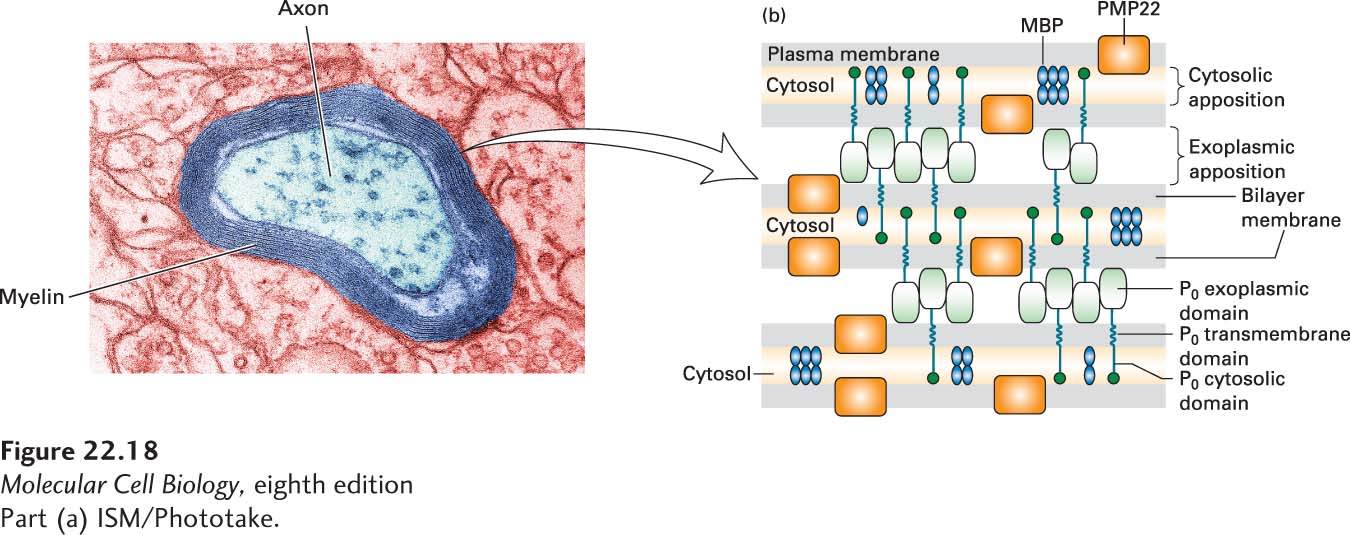
FIGURE 22-18Formation and structure of a myelin sheath in the peripheral nervous system. (a) At high magnification the specialized spiral myelin membrane appears as a series of layers, or lamellae, of phospholipid bilayers wrapped around the axon. (b) Close-up view of three layers of the myelin membrane spiral. The two most abundant integral myelin membrane proteins, P0 and PMP22, are produced only by Schwann cells. The exoplasmic domain of a P0 protein, which has an immunoglobulin fold, associates with similar domains emanating from P0 proteins in the opposite membrane surface, thereby “zippering” together the exoplasmic membrane surfaces in close apposition. These interactions are stabilized by binding of a tryptophan residue on the tip of the exoplasmic domain to lipids in the opposite membrane. Close apposition of the cytosolic faces of the membrane may result from binding of the cytosolic tail of each P0 protein to phospholipids in the opposite membrane. PMP22 may also contribute to membrane compaction. Myelin basic protein (MBP), a cytosolic protein, remains between the closely apposed membranes as the cytosol is squeezed out. See L. Shapiro et al., 1996, Neuron17:435, and E. J. Arroyo and S. S. Scherer, 2000, Histochem. Cell Biol.113:1.
[Part (a) ISM/Phototake.]
Page 1045
Damage to proteins produced by oligodendrocytes underlies a prevalent human neurological disease, multiple sclerosis (MS). MS is usually characterized by spasms and weakness in one or more limbs, bladder dysfunction, local sensory losses, and visual disturbances. This disorder—the prototype demyelinating disease—is caused by patchy loss of myelin in areas of the brain and spinal cord. In MS patients, conduction of action potentials by the demyelinated neurons is slowed, and the Na+ channels spread outward from the nodes, lowering their nodal concentration. The cause of the disease is not known but appears to involve either the body’s production of auto-antibodies (antibodies that bind to normal body proteins) that react with MBP, or the secretion of proteases that destroy myelin proteins. A mouse mutant, shiverer, has a deletion of much of the MBP gene, leading to tremors, convulsions, and early death. Similarly, human (Pelizaeus–Merzbacher disease) and mouse (jimpy) mutations in the gene coding for the other major protein of CNS myelin, PLP, cause loss of oligodendrocytes and inadequate myelination.
Schwann Cells Schwann cells form myelin sheaths around peripheral nerves. A Schwann cell myelin sheath is a remarkable spiral wrap (Figure 22-17b). A long axon can have as many as several hundred Schwann cells along its length, each contributing myelin insulation to an internode stretch of about 1–1.5 µm of axon. For reasons that are not understood, not all axons are myelinated. Mutations in mice that eliminate Schwann cells cause the death of most neurons.
In contrast to oligodendrocytes, each Schwann cell myelinates only one axon. The sheaths are composed of about 70 percent lipid (rich in cholesterol) and 30 percent protein. In the PNS, the principal protein constituent (~80 percent) of myelin is called protein 0 (P0), an integral membrane protein that has immunoglobulin (Ig) domains. MBP is also an abundant component. The extracellular Ig domains of P0 bind together the surfaces of sequential wraps around the axon to compact the spiral of myelin sheath (Figure 22-18). Other proteins play this kind of role in the CNS.
In humans, peripheral myelin, like CNS myelin, is a target of autoimmune disease, mainly involving the formation of antibodies against P0. The Guillain-Barré syndrome (GBS), also known as acute inflammatory demyelinating polyneuropathy, is one such disease. GBS is the most common cause of rapid-onset paralysis, occurring at a frequency of one person out of 100,000. The cause is unknown, although it usually follows an acute infectious illness and is thought to involve an immune attack on the peripheral nervous system. The common inherited neurological disorder called Charcot-Marie-Tooth disease, which damages peripheral motor and sensory nerve function, is due to overexpression of the gene that encodes PMP22 protein, another constituent of peripheral nerve myelin.
Interactions between glia and neurons control the placement and spacing of myelin sheaths, and the assembly of nerve-transmission machinery at the nodes of Ranvier. Voltage-gated Na+ channels and Na+/K+ pumps, for example, congregate at the nodes of Ranvier through interactions with cytoskeletal proteins. While the details of the node assembly process are not fully understood, a number of key players have been identified. In the PNS, where the process has been most studied, surface adhesion molecules in the Schwann cell membrane first interact with neuronal surface adhesion molecules. An immunoglobulin cell-adhesion molecule (IgCAM) in the glial membrane, called neurofascin155, contacts two axonal proteins, contactin and contactin-associated protein, at the edge of the node. These cell-cell contact events create boundaries at each side of the node.
Page 1046
The channel proteins and other molecules that will accumulate at the node are initially dispersed throughout the axons. Then axonal proteins, including two IgCAMs called NrCAM and neurofascin186, as well as ankyrin G (Chapter 17), accumulate within the node. The two IgCAMs bind to a single transmembrane domain protein called gliomedin that is expressed in the glial cell. Experiments that eliminated gliomedin production showed that without it nodes do not form, so it is a key regulator, and demonstrates the importance of glial-neuron interactions in proper development of the nervous system. Ankyrin in the node contacts βIV spectrin, a major constituent of the cytoskeleton, thus tethering the node’s protein complex to the cytoskeleton. Na+ channels become associated with neurofascin186, NrCAM, and ankyrin G, firmly trapping the channel in the nodal segment of the axonal plasma membrane where it is needed. As a result of these multiple protein-protein interactions, the concentration of Na+ channels is roughly a hundredfold higher in the nodal membrane of myelinated axons than in the axonal membrane of nonmyelinated neurons.