B-Cell Development Requires Input from a Pre-B-Cell Receptor
As we have seen, B cells destined to make immunoglobulins must rearrange the necessary gene segments to assemble functional heavy-chain and light-chain genes. These rearrangements occur in a carefully ordered sequence during the development of a B cell, starting with heavy-chain rearrangements. Moreover, the rearranged heavy chain is first used to build a membrane-bound receptor that executes a cell fate decision necessary to drive further B-cell development (and antibody synthesis) by permitting subsequent rearrangement of the light-chain genes. Only a productive rearrangement that yields an in-frame VDJ combination can generate a complete µ heavy chain. The production of that µ chain serves as a signal to the B cell that it has successfully accomplished rearrangement, and that no further rearrangements of the heavy-chain locus on the remaining gene copy are required. Recall that each lymphocyte precursor starts out with two immunoglobulin locus–bearing, homologous chromosomes in the germ-line (unrearranged) configuration. In accordance with clonal selection theory, which stipulates that each lymphocyte ought to come equipped with a single antigen-specific receptor, continued rearrangement would entail the risk of producing B cells with two different heavy chains, each with different specificity—an undesirable outcome.
Successful rearrangement of V, D, and J segments in the heavy-chain locus thus allows the synthesis of a complete µ chain. B cells at this stage of development are called pre-B cells, as they have not yet completed assembly of a functional light-chain gene and therefore cannot engage in antigen recognition. The µ chain is synthesized in the endoplasmic reticulum and becomes part of a membrane-bound signaling receptor whose expression is essential for B-cell development to proceed in orderly fashion.
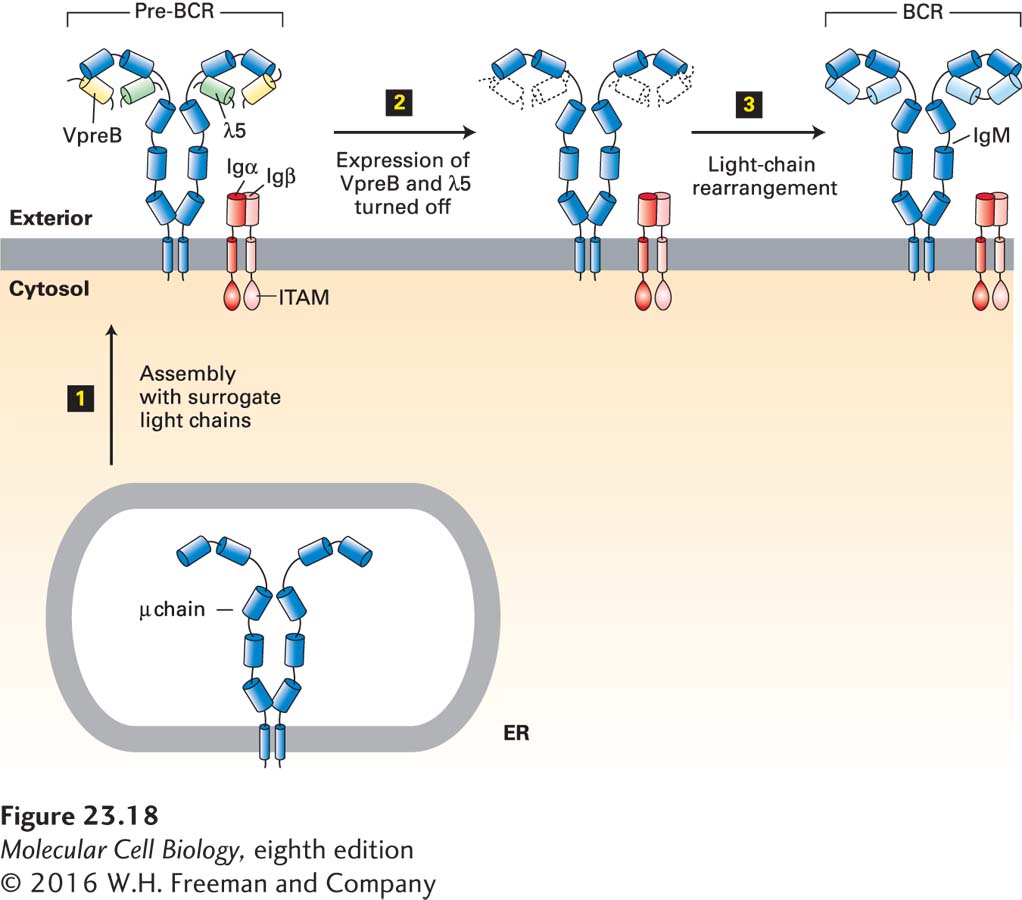
In pre-B cells, newly made µ chains form a complex with two so-called surrogate light chains, λ5 and VpreB (Figure 23-18). The µ chain itself possesses no cytoplasmic tail and is therefore incapable of recruiting cytoplasmic components for the purpose of signal transduction. Instead, pre-B cells express two auxiliary transmembrane proteins, called Igα and Igβ, each of which carries in its cytoplasmic tail an immunoreceptor tyrosine-based activation motif, or ITAM. The entire complex, including µ chain, λ5, VpreB, Igα, and Igβ, constitutes the pre-B-cell receptor (pre-BCR). Engagement of this receptor by (unknown) suitable signals results in recruitment and activation of an Src-family tyrosine kinase, which phosphorylates tyrosine residues in the ITAMs. In their phosphorylated form, ITAMs recruit other molecules essential for signal transduction (see below). Because no functional light chains are yet part of this receptor, it is presumed to be incapable of antigen recognition, as the antigen-binding site has contributions from both the heavy and light chains (see Figure 23-14) (except in camelids).
FIGURE 23-18Structure of the pre-B-cell receptor and its role in B-cell development. Successful rearrangement of V, D, and J heavy-chain gene segments allows synthesis of membrane-bound µ heavy chains in the endoplasmic reticulum (ER) of a pre-B cell. At this stage, no light-chain gene rearrangement has occurred. Newly made µ chains assemble with surrogate light chains, composed of λ5 and VpreB, and Igα/Igβ to yield the pre-B-cell receptor, pre-BCR (step 1). This receptor drives proliferation of those B cells that carry it. It also suppresses rearrangement of the heavy-chain locus on the other chromosome and so mediates allelic exclusion. In the course of proliferation, the synthesis of λ5 and VpreB is shut off (step 2), resulting in “dilution” of the available surrogate light chains and reduced expression of the pre-BCR. As a result, rearrangement of the light-chain loci can proceed (step 3). If this rearrangement is productive, the B cell can synthesize light chains and complete assembly of the B-cell receptor (BCR), which consists of a membrane-bound IgM and associated Igα and Igβ. The B cell is now responsive to antigen-specific stimulation.
The pre-B-cell receptor has several important functions. First, it shuts off expression of the RAG recombinases, so that rearrangement of the other (allelic) heavy-chain locus cannot proceed. This phenomenon, called allelic exclusion, ensures that only one of the two available copies of the heavy-chain locus will be rearranged and thus expressed as a complete µ chain. Second, because of the association of the pre-B-cell receptor with Igα and Igβ, the receptor becomes a functional signal-transduction unit. Signals that emanate from the pre-BCR initiate proliferation of the pre-B-cell to expand the numbers of those B cells that have undergone productive D-J and V-D-J recombination.
In the course of this expansion, expression of the surrogate light chains, VpreB and λ5, subsides. The progressive dilution of VpreB and λ5 with every successive cell division results in insufficient fully assembled pre-BCR in the endoplasmic reticulum. As a consequence, the heavy chains are degraded (see Chapters 13 and 14) and the amount of pre-BCR signaling decreases. This reduction in signaling allows re-initiation of expression of the RAG recombinases, which now target the κ or λ light-chain locus. A productive light-chain V-J rearrangement also shuts off rearrangement of the allelic locus (allelic exclusion). Upon completion of a successful V-J light-chain rearrangement, the B cell can make both µ heavy chains and κ or λ light chains and assemble them into a functional B-cell receptor (BCR), which can recognize antigen (see Figure 23-18).
Page 1101
Once a B cell expresses a complete BCR on its cell surface, it can recognize antigen, and all subsequent steps in B-cell activation and differentiation require engagement with the antigen for which that BCR is specific. The BCR not only plays a role in driving B-cell proliferation upon a successful encounter with antigen, but also functions as a device for receptor-mediated endocytosis, an essential step that allows the B cell to process the acquired antigen and convert it into a signal that sends out a call for assistance by T lymphocytes. This antigen-presentation function of B cells is described in later sections.