Amino Acids Differing Only in Their Side Chains Compose Proteins
The monomeric building blocks of proteins are 20 amino acids, which—when incorporated into a protein polymer—are sometimes called residues. All amino acids have a characteristic structure consisting of a central alpha carbon atom (Cα) bonded to four different chemical groups: an amino (–NH2) group, a carboxyl or carboxylic acid (–COOH) group (hence the name amino acid), a hydrogen (H) atom, and one variable group, called a side chain or R group. Because the α carbon in all amino acids except glycine is asymmetric, these molecules can exist in two mirror-image forms, called by convention the D (dextro) and the L (levo) isomers (see Figure 2-4). The two isomers cannot be interconverted (one made identical to the other) without breaking and then re-forming a chemical bond in one of them. With rare exceptions, only the L forms of amino acids are found in proteins. However, D amino acids are prevalent in bacterial cell walls and other microbial products.
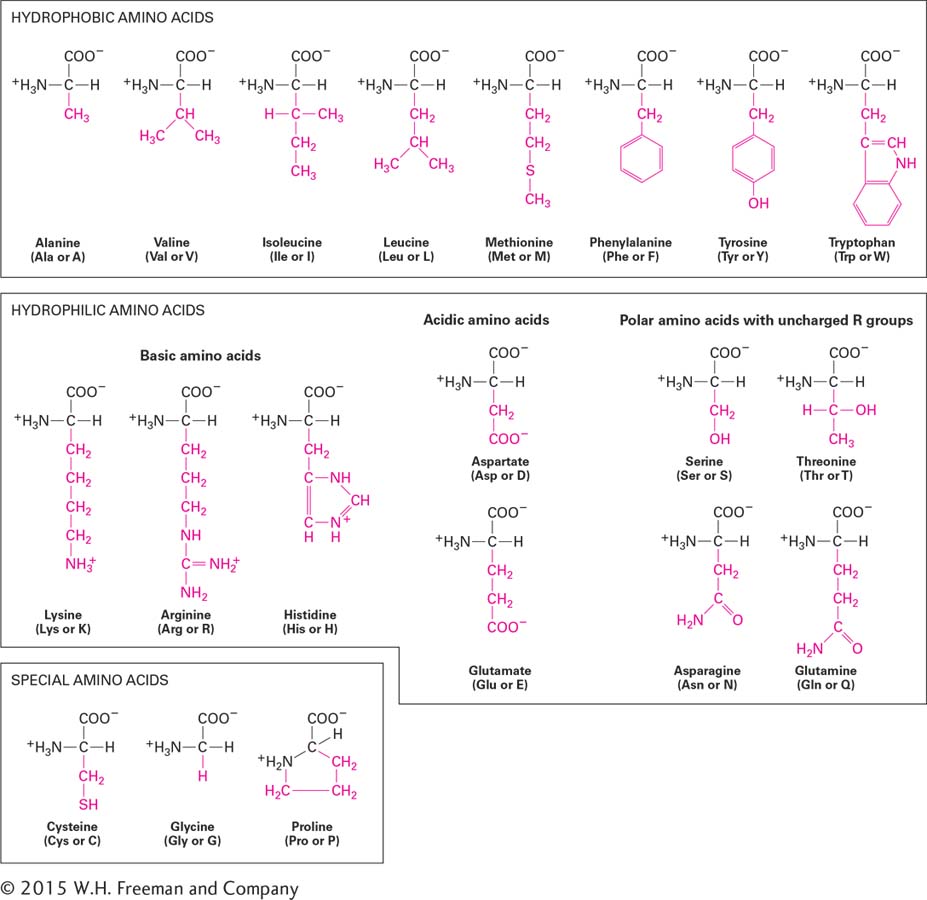
To understand the three-dimensional structures and functions of proteins, discussed in detail in Chapter 3, you must be familiar with some of the distinctive properties of amino acids, which are determined in part by their side chains. You need not memorize the detailed structure of each type of side chain to understand how proteins work because amino acids can be classified into several broad categories based on the size, shape, charge, hydrophobicity (a measure of water solubility), and chemical reactivity of their side chains (Figure 2-14).
Page 43
FIGURE 2-14The 20 common amino acids used to build proteins. The side chain (R group; red) determines the characteristic properties of each amino acid and is the basis for grouping amino acids into three main categories: hydrophobic, hydrophilic, and special. Shown are the ionized forms that exist at the pH (∼7) of the cytosol. In parentheses are the three-letter and one-letter abbreviations for each amino acid.
Amino acids with nonpolar side chains, called hydrophobic amino acids, are poorly soluble in water. The larger the nonpolar side chain, the more hydrophobic the amino acid. The side chains of alanine, valine, leucine, and isoleucine are linear or branched hydrocarbons that do not form a ring, and they are therefore called aliphatic amino acids. These amino acids are all nonpolar, as is methionine, which is similar to them except that it contains one sulfur atom. Phenylalanine, tyrosine, and tryptophan have large, hydrophobic, aromatic rings in their side chains. In later chapters, we will see in detail how hydrophobic side chains under the influence of the hydrophobic effect often pack into the interior of proteins or line the surfaces of proteins that are embedded within hydrophobic regions of biomembranes.
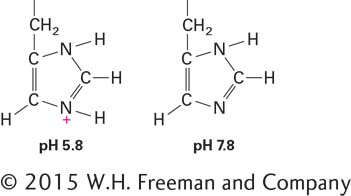
Amino acids with polar side chains are called hydrophilic amino acids; the most hydrophilic of these amino acids is the subset with side chains that are charged (ionized) at the pH typical of biological fluids (∼7) both inside and outside the cell (see Section 2.3). Arginine and lysine have positively charged side chains and are called basic amino acids; aspartic acid and glutamic acid have negatively charged side chains due to the carboxylic acid groups in their side chains (their charged forms are called aspartate and glutamate) and are called acidic amino acids. A fifth amino acid, histidine, has a side chain containing a ring with two nitrogens, called imidazole, which can shift from being positively charged to uncharged in response to small changes in the acidity of its environment:
Page 44
The activities of many proteins are modulated by shifts in environmental acidity (pH) through protonation or deprotonation of histidine side chains. Asparagine and glutamine are uncharged but have polar side chains containing amide groups with extensive hydrogen-bonding capacities. Similarly, serine and threonine are uncharged but have polar hydroxyl groups, which also participate in hydrogen bonds with other polar molecules.
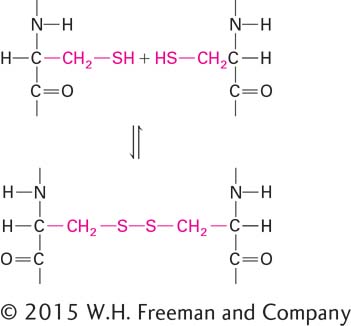
Finally, cysteine, glycine, and proline play special roles in proteins because of the unique properties of their side chains. The side chain of cysteine contains a reactive sulfhydryl group (–SH). On release of a proton (H+), a sulfhydryl group is converted into a thiolate anion (S–). Thiolate anions can play important roles in catalysis, notably in certain enzymes that destroy proteins (proteases). In proteins, each of two adjacent sulfhydryl groups can be oxidized, each releasing a proton and an electron, to form a covalent disulfide bond (–S–S–):
Disulfide bonds serve to “cross-link” regions within a single polypeptide chain (intramolecular cross-linking) or between two separate chains (intermolecular cross-linking). Disulfide bonds stabilize the folded structure of some proteins. The smallest amino acid, glycine, has a single hydrogen atom as its R group. Its small size allows it to fit into tight spaces. Unlike those of the other common amino acids, the side chain of proline (pronounced pro-leen) bends around to form a ring by covalently bonding to the nitrogen atom in the amino group attached to the Cα. As a result, proline is very rigid, and its amino group is not available for typical hydrogen bonding. The presence of proline in a protein creates a fixed kink in the polymer chain, limiting how it can fold in the vicinity of the proline residue.
Some amino acids are more abundant in proteins than others. Cysteine, tryptophan, and methionine are not common amino acids: together, they constitute approximately 5 percent of the amino acids in a typical protein. Four amino acids—leucine, serine, lysine, and glutamic acid—are the most abundant amino acids, constituting 32 percent of all the residues in a typical protein. However, the amino acid compositions of particular proteins may vary widely from these values.
Humans and other mammals can synthesize 11 of the 20 amino acids. The other nine are called essential amino acids and must be included in the diet to permit normal protein production. These essential amino acids are phenylalanine, valine, threonine, tryptophan, isoleucine, methionine, leucine, lysine, and histidine. Adequate provision of these essential amino acids in feed is key to the livestock industry. Indeed, a genetically engineered variety of corn with a high lysine content is now in use as an “enhanced” feed to promote the growth of animals.
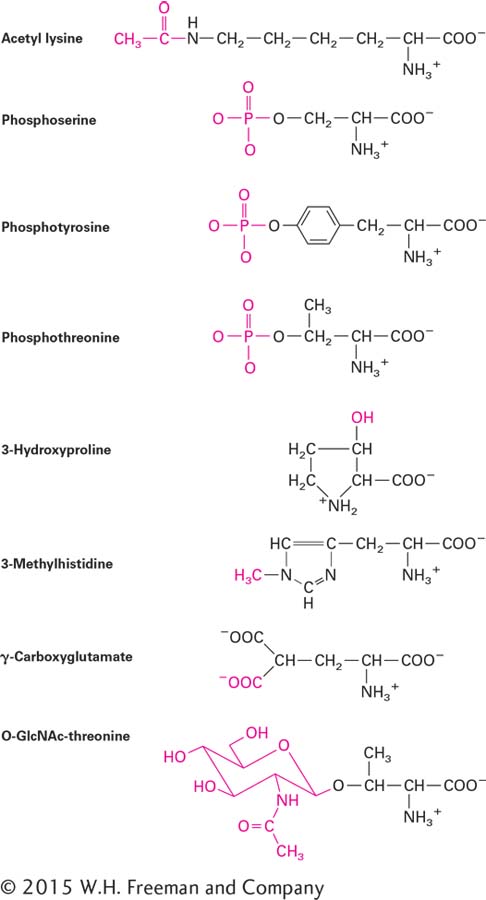
Although cells use the 20 amino acids shown in Figure 2-14 in the initial synthesis of proteins, analysis of cellular proteins reveals that they contain over 100 different amino acids. The difference is due to the chemical modification of some amino acids after they are incorporated into proteins by the addition of a variety of chemical groups (Figure 2-15). One important modification is the addition of acetyl groups (CH3CO) to amino acids, a process known as acetylation. Another is the addition of a phosphate (PO4) to hydroxyl groups in serine, threonine, and tyrosine residues, a process known as phosphorylation. We will encounter numerous examples of proteins whose activity is regulated by reversible phosphorylation and dephosphorylation. Phosphorylation of nitrogen in the side chain of histidine is well known in bacteria, fungi, and plants, but less studied—perhaps because of the relative instability of phosphorylated histidine—and apparently rare in mammals. Methylation of arginine and lysine side chains on proteins called histones is an important regulator of gene expression in eukaryotes (see Chapter 9). Like phosphorylation and dephosphorylation, controlled methylation and demethylation are important regulatory processes. The side chains of asparagine, serine, and threonine are sites for glycosylation, the attachment of linear and branched carbohydrate chains. Many secreted proteins and membrane proteins contain glycosylated residues, and the reversible modification of hydroxyl groups on specific serines and threonines by a sugar called N-acetylglucosamine also regulates protein activities. Other amino acid modifications found in selected proteins include the hydroxylation of proline and lysine residues in collagen (see Chapter 19), the methylation of histidine residues in membrane receptors, and the γ-carboxylation of glutamate in blood-clotting factors such as prothrombin. Deamidation of asparagine and glutamine to form the corresponding acidic amino acids, aspartate and glutamate, is also a common occurrence.
Page 45
FIGURE 2-15Common modifications of amino acid side chains in proteins. These modified residues and numerous others are formed by addition of various chemical groups (red) to the amino acid side chains during or after synthesis of a polypeptide chain.
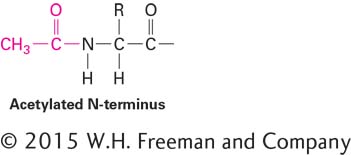
Acetylation of the amino group of the N-terminal residue is the most common form of amino acid chemical modification, affecting an estimated 80 percent of all proteins:
This modification may play an important role in controlling the life span of proteins within cells because many nonacetylated proteins are rapidly degraded.