Multiple Polypeptides Assemble into Quaternary Structures and Supramolecular Complexes
Multimeric proteins consist of two or more polypeptide chains, which in this context are referred to as subunits. A fourth level of structural organization, quaternary structure, describes the number (stoichiometry) and relative positions of the subunits in multimeric proteins (Figure 3-2). Flu virus hemagglutinin, for example, is a trimer of three identical subunits (a homotrimer) held together by noncovalent bonds (Figure 3-11b). Other multimeric proteins are composed of various numbers of identical (homomeric) or different (heteromeric) subunits. Hemoglobin, the oxygen-carrying molecule in blood, is an example of a heteromeric multimeric protein, as it has two copies each of two different polypeptide chains (as discussed below). In many cases, the individual monomer subunits of a multimeric protein cannot function normally unless they are assembled into the multimeric protein. In other cases, assembly into a multimeric protein permits proteins that act sequentially in a pathway to increase their efficiency of operation owing to their juxtaposition in space, a phenomenon referred to as metabolic coupling. Classic examples of metabolic coupling are the fatty acid synthases, the enzymes in fungi that synthesize fatty acids, and the polyketide synthases, the large multiprotein complexes in bacteria that synthesize a diverse set of pharmacologically relevant molecules called polyketides, including the antibiotic erythromycin.
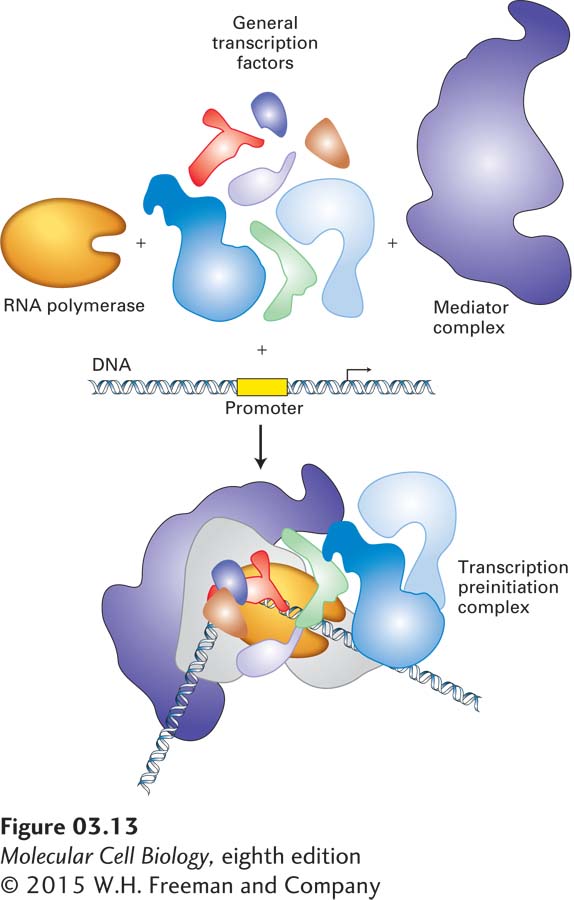
The highest level in the hierarchy of protein structure is the association of proteins into supramolecular complexes. Typically, such structures are very large, in some cases exceeding 1 megadalton (MDa) in mass, approaching 30–300 nm in size, and containing tens to hundreds of polypeptide chains and sometimes other biopolymers such as nucleic acids. The capsid that encases the nucleic acids of the viral genome is an example of a supramolecular complex with a structural function. The bundles of cytoskeletal filaments that support and give shape to the plasma membrane are another example. Other supramolecular complexes act as molecular machines, carrying out the most complex cellular processes by integrating multiple proteins, each with distinct functions, into one large assembly. For example, a transcriptional machine is responsible for synthesizing messenger RNA (mRNA) using a DNA template. This transcriptional machine, the operational details of which are discussed in Chapters 5 and 9, consists of RNA polymerase, itself a multimeric protein, and at least 50 additional components, including general transcription factors, promoter-binding proteins, helicase, and other protein complexes (Figure 3-13). Ribosomes, also discussed in Chapter 5, are complex multiprotein and multi-nucleic acid machines that synthesize proteins. One of the most complex multiprotein assemblies is the nuclear pore, a structure that allows communication and passage of macromolecules between the nucleus and the cytoplasm (see Chapter 14). It is composed of multiple copies of about 30 distinct proteins and forms an assembly with an estimated mass of 50 MDa. The fatty acid synthases and polyketide synthases referred to above are also molecular machines.
Page 79
FIGURE 3-13A molecular machine: the transcription initiation complex. The core RNA polymerase, general transcription factors, a mediator complex containing about 20 subunits, and other protein complexes not depicted here assemble at a promoter in DNA. The polymerase carries out transcription of DNA; the associated proteins are required for initial binding of the polymerase to a specific promoter. The multiple components function together as a molecular machine.