Phospholipid Bilayers Form a Sealed Compartment Surrounding an Internal Aqueous Space
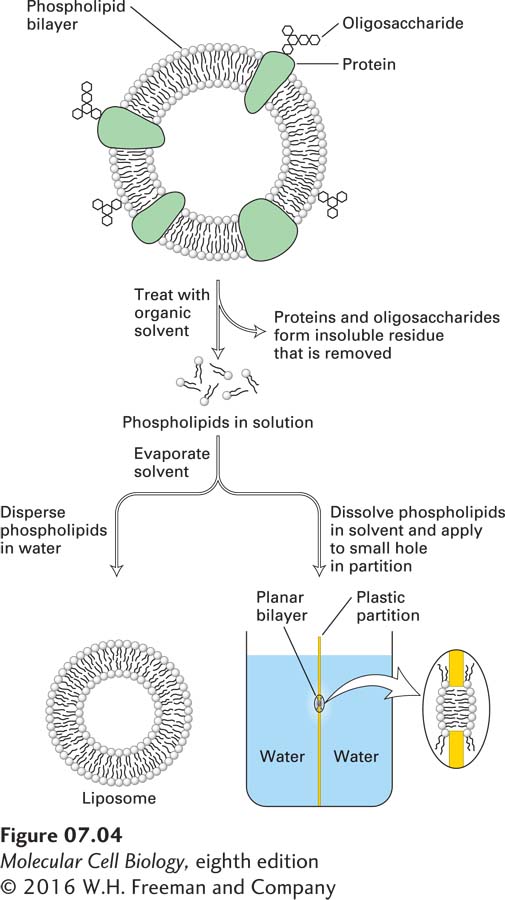
Phospholipid bilayers can be generated in the laboratory by simple means, using either chemically pure phospholipids or lipid mixtures of the composition found in cellular membranes (Figure 7-4). Such synthetic bilayers possess three important properties. First, they are virtually impermeable to water-soluble (hydrophilic) solutes, which do not readily diffuse across the bilayer. These solutes include salts, sugars, and most other small hydrophilic molecules—including water itself. The second property of a bilayer is its stability. The hydrophobic and van der Waals interactions between the fatty acyl chains maintain the integrity of the interior of the bilayer structure. Even though the exterior aqueous environment may vary widely in ionic strength and pH, the bilayer has the strength to retain its characteristic architecture. Third, all synthetic phospholipid bilayers can spontaneously form sealed closed compartments in which the aqueous space on the inside is separated from the exterior environment. An “edge” of a phospholipid bilayer, as depicted in Figure 7-3b, with the hydrocarbon core of the bilayer exposed to an aqueous solution, would be unstable; the exposed fatty acyl chains would be in an energetically much more stable state if they were not adjacent to water molecules but rather surrounded by other fatty acyl chains. Thus, in an aqueous solution, sheets of phospholipid bilayers spontaneously seal their edges, forming a spherical bilayer that encloses an aqueous central compartment. The liposome depicted in Figure 7-3c is an example of such a structure viewed in cross section.
EXPERIMENTAL FIGURE 7-4Formation and study of pure phospholipid bilayers. (Top) A preparation of biological membranes is treated with an organic solvent, such as a mixture of chloroform and methanol (3:1), which selectively solubilizes the phospholipids and cholesterol. Proteins and carbohydrates remain in an insoluble residue. The solvent is removed by evaporation. (Bottom left) If the extracted materials are mechanically dispersed in water, they spontaneously form a liposome, shown in cross section, with an internal aqueous compartment. (Bottom right) A planar bilayer, also shown in cross section, can form over a small hole in a partition separating two aqueous phases; such a system can be used to study the physical properties of bilayers, such as their permeability to solutes.
Page 275
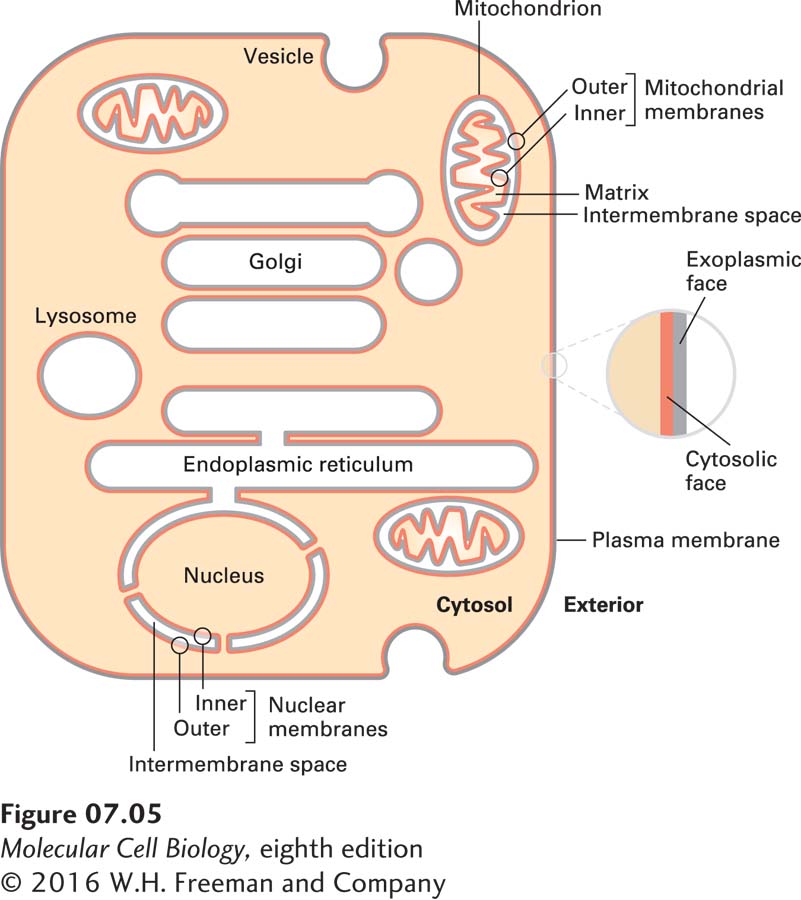
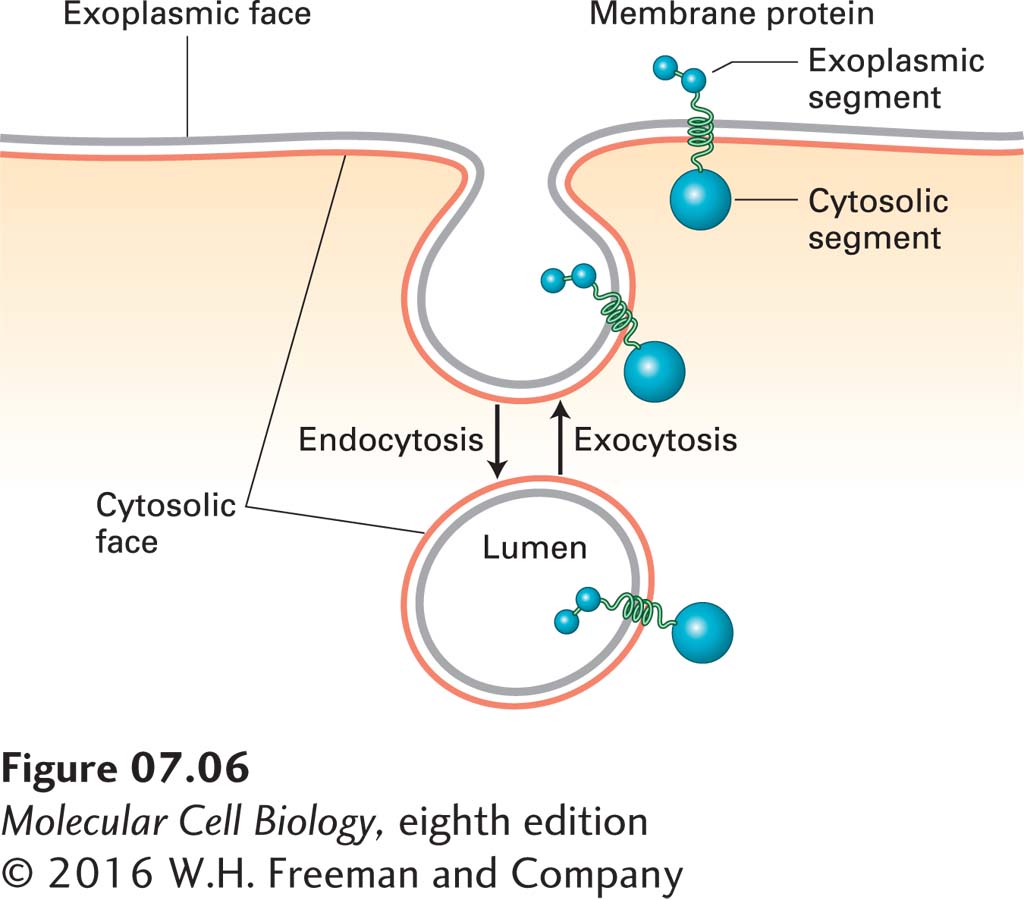
This physical chemical property of a phospholipid bilayer has important implications for cellular membranes: no membrane in a cell can have an “edge” with exposed hydrocarbon fatty acyl chains. All membranes form closed compartments, similar in basic architecture to liposomes. Because all cellular membranes enclose an entire cell or an internal compartment, they have an internal face (the surface oriented toward the interior of the compartment) and an external face (the surface presented to the environment). More commonly, we designate the two surfaces of a cellular membrane as the cytosolic face and the exoplasmic face. This nomenclature is useful in highlighting the topological equivalence of the faces in different membranes, as diagrammed in Figures 7-5 and 7-6. For example, the exoplasmic face of the plasma membrane is directed away from the cytosol, toward the extracellular space or external environment, and defines the outer limit of the cell. The cytosolic face of the plasma membrane faces the cytosol. Similarly for organelles and vesicles surrounded by a single membrane, the cytosolic face faces the cytosol. The exoplasmic face is always directed away from the cytosol, and in this case it is on the inside of the organelle, in contact with the internal aqueous space, or lumen. The lumen of a vesicle is topologically equivalent to the extracellular space, a concept most easily understood for vesicles that arise by invagination (endocytosis) of the plasma membrane. The external face of the plasma membrane becomes the internal face of the vesicle membrane, while in the vesicle the cytosolic face of the plasma membrane still faces the cytosol (see Figure 7-6).
FIGURE 7-5The faces of cellular membranes. The plasma membrane, a single bilayer, encloses the cell. In this highly schematic representation, internal cytosol (tan) and external environment (white) define the cytosolic (red) and exoplasmic (gray) faces of the bilayer. Vesicles and some organelles have a single membrane, and their internal aqueous space (white) is topologically equivalent to the outside of the cell. Three organelles—the nucleus, mitochondrion, and chloroplast (which is not shown)—are enclosed by two membranes separated by a small intermembrane space. The exoplasmic faces of the inner and outer membranes around these organelles border the intermembrane space between them. For simplicity, the hydrophobic membrane interior is not indicated in this diagram.
FIGURE 7-6The faces of cellular membranes are conserved during membrane budding and fusion. Red membrane surfaces are cytosolic faces; gray membrane surfaces are exoplasmic faces. During endocytosis, a segment of the plasma membrane buds inward toward the cytosol and eventually pinches off a separate vesicle. During this process, the cytosolic face of the plasma membrane remains facing the cytosol, and the exoplasmic face of the new vesicle membrane faces the vesicle lumen. During exocytosis, an intracellular vesicle fuses with the plasma membrane, and the lumen of the vesicle (exoplasmic face) connects with the extracellular medium. Proteins that span the membrane retain their asymmetric orientation during vesicle budding and fusion; in particular, the same segment always faces the cytosol.
Three organelles—the nucleus, mitochondrion, and chloroplast—are surrounded not by a single membrane, but by two. The exoplasmic surface of each membrane faces the space between the two membranes (the intermembrane space). This relationship can perhaps best be understood by reference to the endosymbiont hypothesis, discussed in Chapter 12, which posits that mitochondria and chloroplasts arose early in the evolution of eukaryotic cells by the engulfment of bacteria capable of oxidative phosphorylation or photosynthesis, respectively (see Figure 12-7).
Page 276
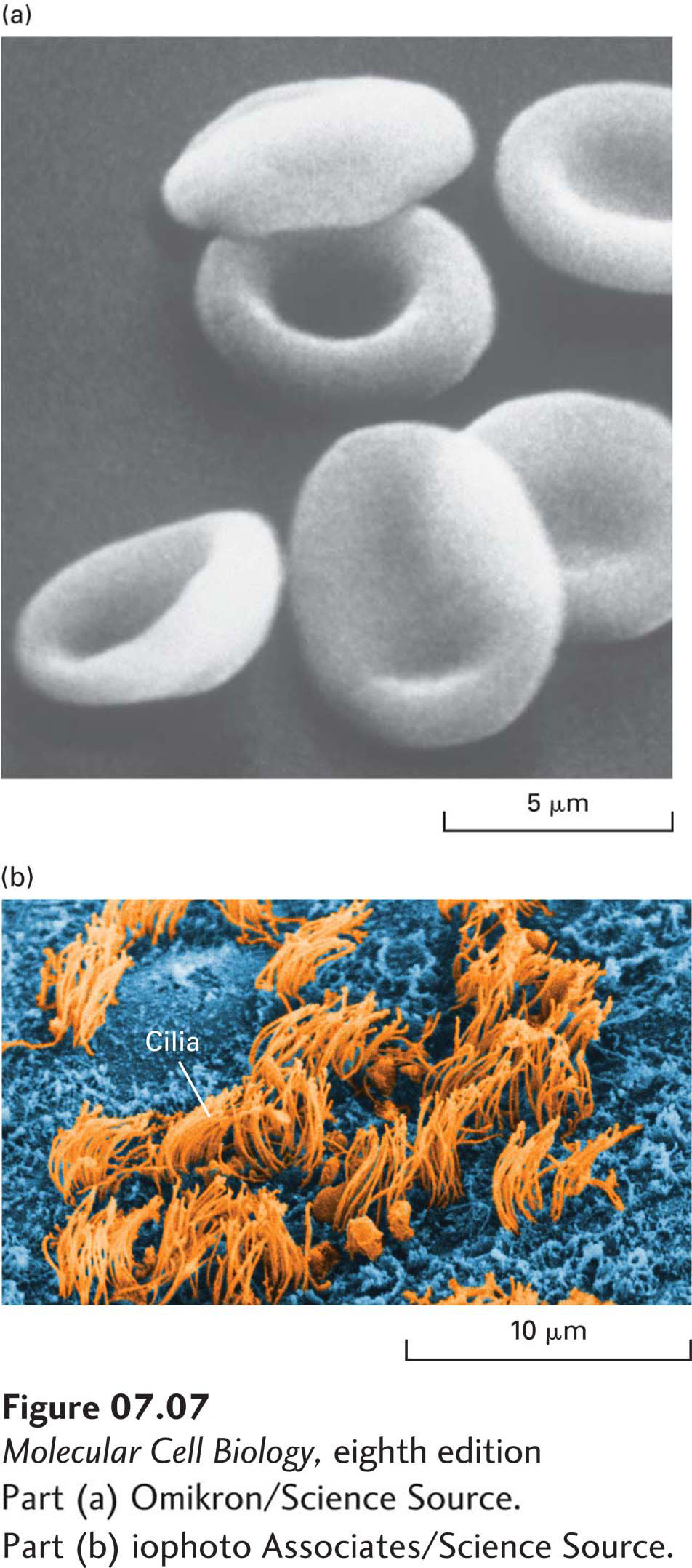
Natural membranes from different cell types exhibit a variety of shapes, which complement a cell’s function. The smooth, flexible surface of the erythrocyte plasma membrane allows the cell to squeeze through narrow blood capillaries (Figure 7-7a). Some cells have a long, slender extension of the plasma membrane, called a cilium or flagellum, which beats in a whiplike manner (Figure 7-7b). This motion causes fluid to flow across the surface of a sheet of cells, or a sperm cell to swim toward an egg. The differing shapes and properties of biomembranes raise a key question in cell biology, namely, how the composition of biological membranes is regulated to establish and maintain the identity of the different membrane structures and membrane-delimited compartments. We return to this question in Section 7.3 and in Chapter 14.
[Part (a) Omikron/Science Source. Part (b) iophoto Associates/Science Source.]
FIGURE 7-7Variation in biomembranes in different cell types. (a) A smooth, flexible membrane covers the surface of the discoid erythrocyte cell, as seen in this scanning electron micrograph. (b) Tufts of cilia project from the ependymal cells that line the brain ventricles.
[Part (a) Omikron/Science Source. Part (b) iophoto Associates/Science Source.]