Lipid Composition Is Different in the Exoplasmic and Cytosolic Leaflets
A characteristic of all biomembranes is an asymmetry in lipid composition across the bilayer. Although most phospholipids are present in both membrane leaflets, some are commonly more abundant in one or the other leaflet. For instance, in plasma membranes from human erythrocytes and Madin-Darby canine kidney (MDCK) cells grown in culture, almost all the sphingomyelin and phosphatidylcholine, both of which form less fluid bilayers, are found in the exoplasmic leaflet. In contrast, phosphatidylethanolamine, phosphatidylserine, and phosphatidylinositol, which form more fluid bilayers, are preferentially located in the cytosolic leaflet. Because phosphatidylserine and phosphatidylinositol carry a net negative charge, the stretch of amino acids on the cytosolic face of a membrane protein with a single transmembrane segment, is often enriched in positively charged (Lys, Arg) residues in close proximity to the transmembrane segment (this distribution of charged amino acids is referred to as the “inside positive” rule). This segregation of lipids across the bilayer may influence membrane curvature (see Figure 7-11c). Unlike particular phospholipids, cholesterol is relatively evenly distributed in both leaflets of cellular membranes.
Page 282
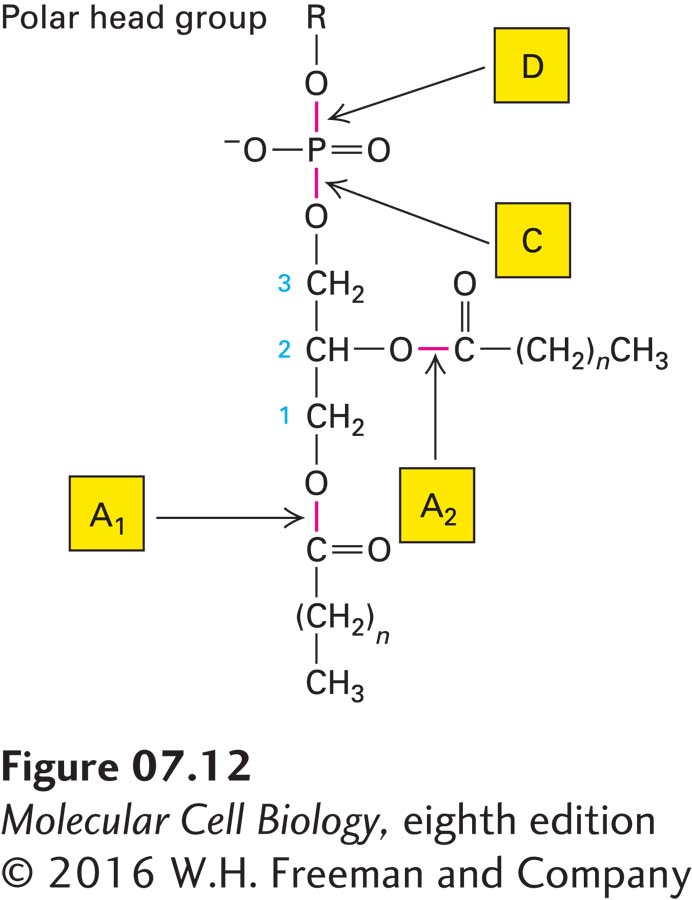
The relative abundances of a particular phospholipid in the two leaflets of a plasma membrane can be determined experimentally on the basis of the susceptibility of phospholipids to hydrolysis by phospholipases, enzymes that cleave the ester bonds via which acyl chains and head groups are connected to the lipid molecule (Figure 7-12). When added to the external medium, phospholipases cannot cross the membrane, and thus they cleave off the head groups of only those lipids present in the exoplasmic face; phospholipids in the cytosolic leaflet are resistant to hydrolysis because the enzymes cannot penetrate to the cytosolic face of the plasma membrane.
FIGURE 7-12Specificity of phospholipases. Each type of phospholipase cleaves one of the susceptible bonds shown in red. The glycerol carbon atoms are indicated by small numbers. In intact cells, only phospholipids in the exoplasmic leaflet of the plasma membrane are cleaved by phospholipases in the surrounding medium. Phospholipase C, a cytosolic enzyme, cleaves certain phospholipids in the cytosolic leaflet of the plasma membrane.
How the asymmetric distribution of phospholipids in membrane leaflets arises is still unclear. As noted, in pure bilayers, phospholipids do not spontaneously migrate, or flip-flop, from one leaflet to the other. In part, the asymmetry in phospholipid distribution may reflect where these lipids are synthesized in the endoplasmic reticulum and Golgi. Sphingomyelin is synthesized on the luminal (exoplasmic) face of the Golgi, which becomes the exoplasmic face of the plasma membrane. In contrast, phosphoglycerides are synthesized on the cytosolic face of the ER membrane, which is topologically equivalent to the cytosolic face of the plasma membrane (see Figure 7-5). Clearly, however, this explanation does not account for the preferential location of phosphatidylcholine (a phosphoglyceride) in the exoplasmic leaflet. Movement of this phosphoglyceride, and perhaps others, from one leaflet to the other in some natural membranes is most likely catalyzed by ATP-powered transport proteins called flippases, which are discussed in Chapter 11.
The preferential location of lipids on one face of the bilayer is necessary for a variety of membrane-based functions. For example, the head groups of all phosphorylated forms of phosphatidylinositol (PI; see Figure 7-8), an important source of second messengers, face the cytosol. Stimulation of many cell-surface receptors by their corresponding ligands results in activation of the cytosolic enzyme phospholipase C, which can then hydrolyze the bond within PI connecting the phosphoinositols to the diacylglycerol. As we will see in Chapter 15, both water-soluble phosphoinositols and membrane-embedded diacylglycerol participate in intracellular signaling pathways that affect many aspects of cellular metabolism. Phosphatidylserine too is normally most abundant in the cytosolic leaflet of the plasma membrane. In the initial stages of platelet stimulation by serum, phosphatidylserine is briefly translocated to the exoplasmic face, presumably by a flippase enzyme, where it activates enzymes participating in blood clotting. When cells die, lipid asymmetry is no longer maintained, and phosphatidylserine, normally enriched in the cytosolic leaflet, is increasingly found in the exoplasmic one. This increased exposure is detected experimentally by use of a labeled version of annexin V, a protein that specifically binds to phosphatidylserine, to measure the onset of programmed cell death (apoptosis). The increased exposure of phosphatidylserine on dying or dead cells is recognized by phagocytic cells, which initiate engulfment of such apoptotic bodies and thus ensure timely and safe disposal of cell remnants.