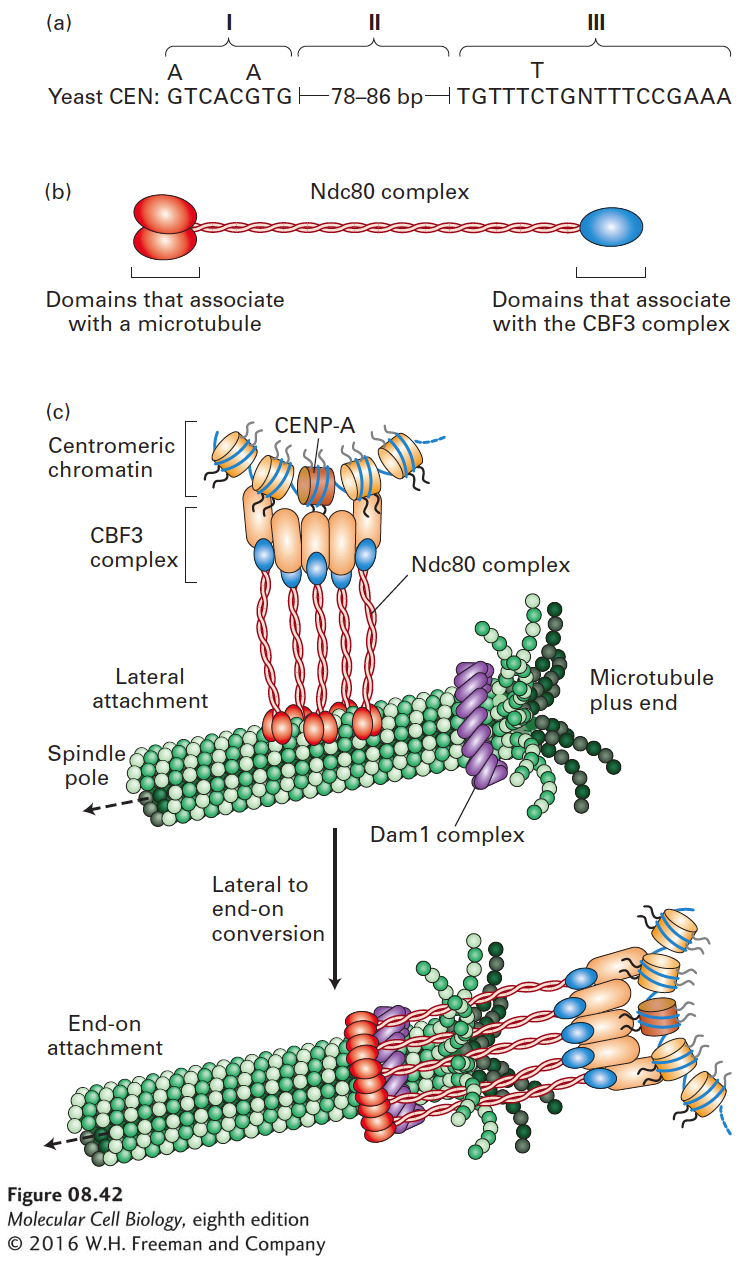
Centromere Sequences Vary Greatly in Length and Complexity
Once the yeast centromere regions that confer mitotic segregation were cloned, their sequences could be determined and compared. The results revealed three regions (I, II, and III) that are conserved among the centromeres on different yeast chromosomes (Figure 8-42a). Short, fairly well-conserved nucleotide sequences are present in regions I and III. Region II does not have a specific sequence, but is AT-rich with a fairly constant length, probably so that regions I and III will lie on the same side of a specialized centromere-associated histone octamer. This specialized centromere-associated histone octamer contains the usual histones H2A, H2B, and H4, but a variant form of histone H3. Centromeres from all eukaryotes similarly contain nucleosomes with a specialized, centromere-specific form of histone H3, called CENP-A in humans. In the simple kinetochore of S. cerevisiae, a protein complex called CBF3 associates with this specialized nucleosome. The CBF3 complex, in turn, associates with several copies of an elongated multiprotein complex called Ndc80 (Figure 8-42b). The Ndc80 complexes initially make lateral interactions with a spindle microtubule and subsequently interact with a Dam1 complex, which forms a ring around the end of the microtubule (Figure 8-42c). This interaction results in an end-on attachment of the centromere to the spindle microtubule. S. cerevisiae has by far the simplest centromere known in nature.
FIGURE 8-42Kinetochore-microtubule interaction in S. cerevisiae. (a) Sequence of the simple centromeres of S. cerevisiae. See L. Clarke and J. Carbon, 1985, Ann. Rev. Genet.19:29. (b) Ndc80 complexes associate with both the microtubule and the CBF3 complex. (c) Diagram of the centromere-associated CBF3 complex and its associated Ndc80 complexes, which associate with a ring of Dam1 proteins at the end of a spindle microtubule. The Ndc80 complexes initially make lateral interactions with the side of a spindle microtubule (top) and then associate with the Dam1 ring, making an end-on attachment (bottom) to the microtubule. See T. U. Tanaka, 2010, EMBO J.29:4070.
Page 346
In the fission yeast S. pombe, centromeres are 40–100 kb in length and are composed of repeated copies of sequences similar to those in S. cerevisiae centromeres. Multiple copies of proteins homologous to those that interact with S. cerevisiae centromeres bind to these complex S. pombe centromeres, and in turn bind the much longer S. pombe chromosomes to several microtubules of the mitotic spindle apparatus. In plants and animals, centromeres are megabases in length and are composed of multiple repeats of simple-sequence DNA. In humans, centromeres contain 2–4-Mb arrays of a 171-bp simple-sequence DNA called alphoid DNA, which is bound by nucleosomes containing the CENP-A histone H3 variant, as well as other repeated simple-sequence DNAs.
Page 347
In higher eukaryotes, a complex protein structure called the kinetochore assembles at centromeres and associates with multiple mitotic spindle fibers during mitosis (see Figure 18-40). Homologs of many of the centromere-associated proteins found in the yeasts occur in humans and other higher eukaryotes. For those yeast proteins for which clear homologs are not evident in higher cells based on amino acid sequence comparisons (such as the Dam1 complex), alternative complexes with similar properties have been proposed to function at kinetochores. The functions of the centromere and of the kinetochore proteins that bind to it during the segregation of sister chromatids in mitosis and meiosis are described in Chapters 18 and 19.