Three Eukaryotic RNA Polymerases Catalyze Formation of Different RNAs
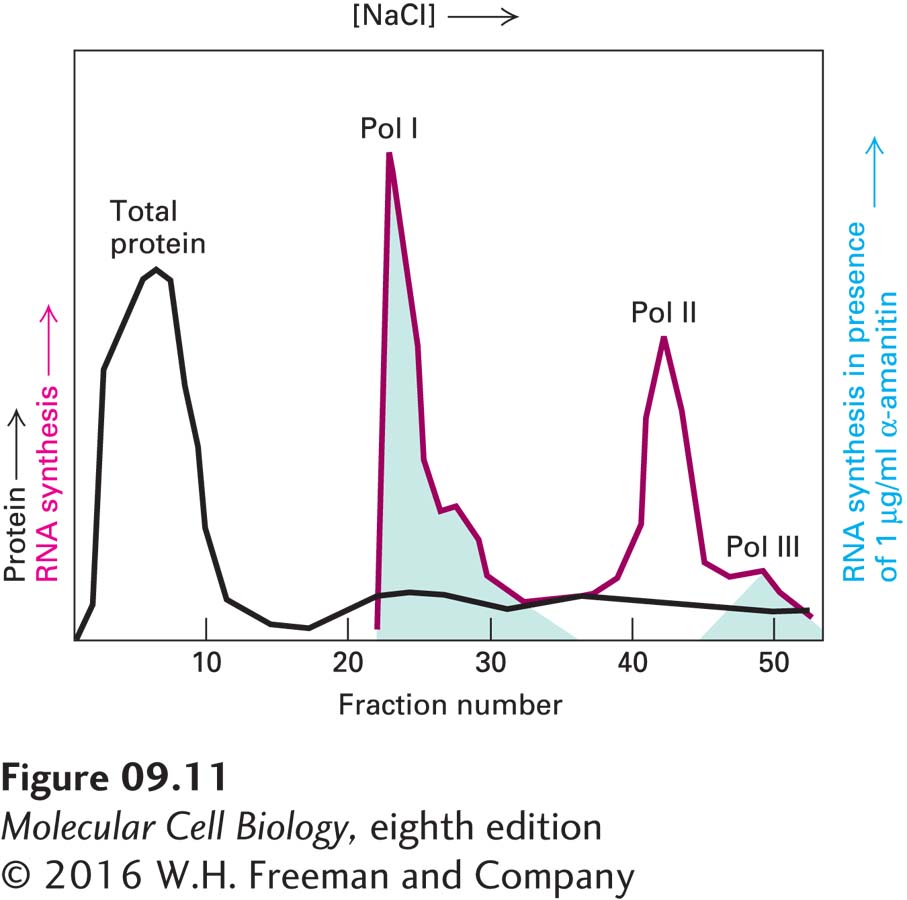
EXPERIMENTAL FIGURE 9-11Liquid chromatography separates and identifies the three eukaryotic RNA polymerases, each with its own sensitivity to α-amanitin. A protein extract from the nuclei of cultured eukaryotic cells was passed through a DEAE Sephadex column and adsorbed protein eluted (black curve) with a solution of constantly increasing NaCl concentration. An aliquot of each fraction of eluate collected from the column was assayed for RNA polymerase activity without (red curve) and with (green shading) 1 µg/ml α-amanitin. This concentration of α-amanitin inhibits polymerase II activity but has no effect on polymerases I and III. Polymerase III is inhibited by 10 µg/ml of α-amanitin, whereas polymerase I is unaffected even at this higher concentration. See R. G. Roeder, 1974, J. Biol. Chem.249:241.
The nuclei of all eukaryotic cells examined so far (e.g., vertebrate, Drosophila, yeast, and plant cells) contain three different RNA polymerases, designated I, II, and III. These enzymes are eluted at different salt concentrations during ion-exchange chromatography, reflecting the differences in their net charges. The three nuclear RNA polymerases also differ in their sensitivity to α-amanitin, a poisonous cyclic octapeptide produced by some mushrooms (Figure 9-11). RNA polymerase I is insensitive to α-amanitin, but RNA polymerase II is very sensitive—the drug binds near the active site of the enzyme and inhibits translocation of the enzyme along the DNA template. RNA polymerase III has intermediate sensitivity.
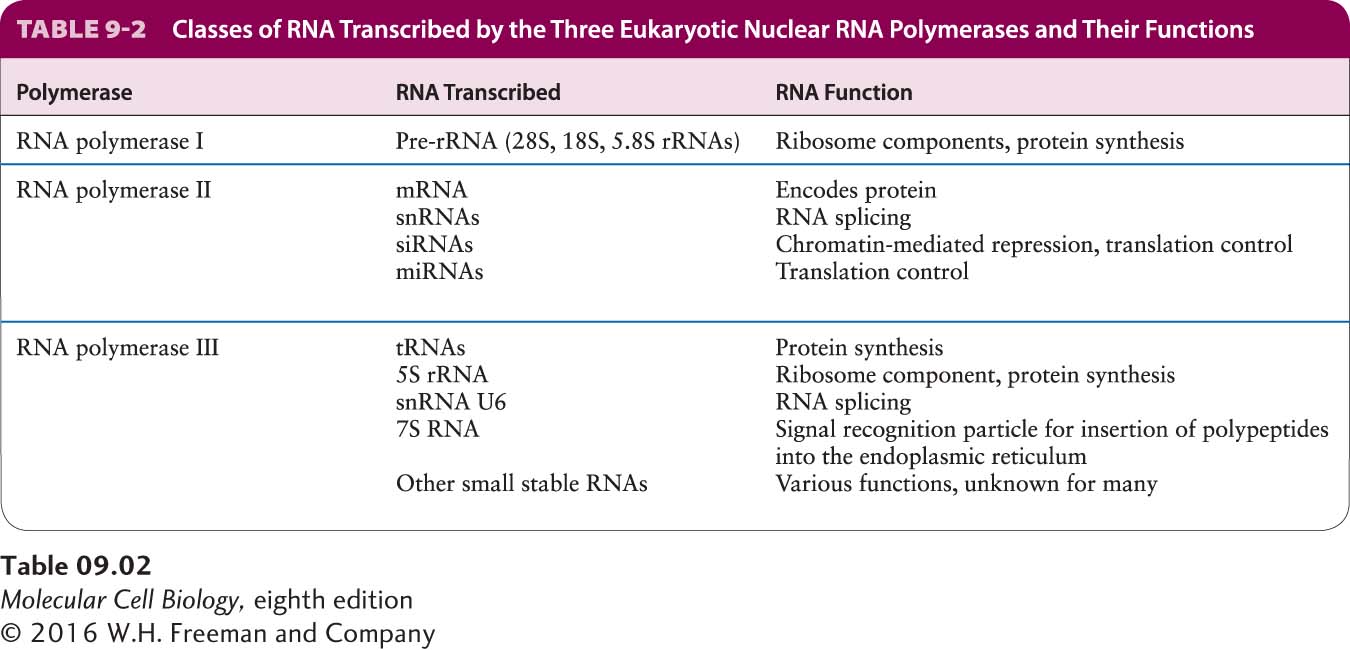
Each eukaryotic RNA polymerase catalyzes transcription of genes encoding different classes of RNA (Table 9-2). RNA polymerase I (Pol I), located in the nucleolus, transcribes genes encoding precursor rRNA (pre-rRNA), which is processed into 28S, 5.8S, and 18S rRNAs. RNA polymerase III (Pol III) transcribes genes encoding tRNAs, 5S rRNA, and an array of small stable RNAs, including one involved in RNA splicing (U6) and the RNA component of the signal recognition particle (SRP) involved in directing nascent proteins to the endoplasmic reticulum (see Chapter 13). RNA polymerase II (Pol II) transcribes all protein-coding genes: that is, it functions in production of mRNAs. RNA polymerase II also produces four of the five small nuclear RNAs (snRNAs) that take part in RNA splicing and micro-RNAs (miRNAs) involved in translation control, as well as the closely related endogenous small interfering RNAs (siRNAs) (see Chapter 10).
Page 368
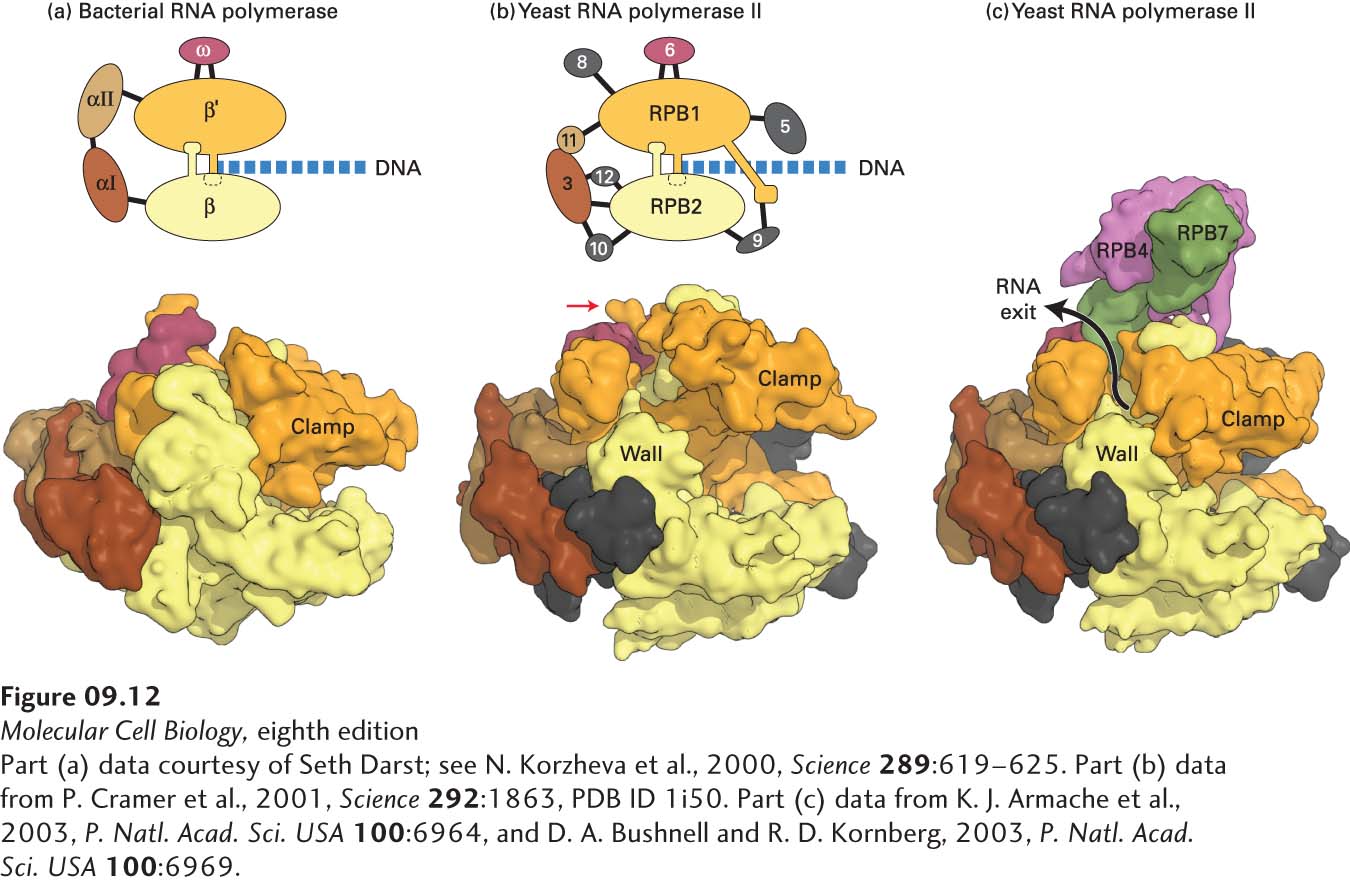
Each of the three eukaryotic RNA polymerases is more complex than E. coli RNA polymerase, but all four of these multisubunit RNA polymerases have a similar overall design (Figure 9-12a, b). All three eukaryotic RNA polymerases contain two large subunits and 10–14 smaller subunits, some of which are common between two or all three of the polymerases. The best-characterized eukaryotic RNA polymerases are from the yeast Saccharomyces cerevisiae. Each of the yeast genes encoding the polymerase subunits has been subjected to gene-knockout mutations and the resulting phenotypes characterized. In addition, the three-dimensional structure of yeast RNA polymerase II has been determined (Figure 9-12b, c). The three nuclear RNA polymerases from all eukaryotes so far examined are very similar to those of yeast. Plants contain two additional nuclear RNA polymerases (RNA polymerases IV and V), which are closely related to their RNA polymerase II but have a unique large subunit and some additional unique subunits. These two polymerases function in transcriptional repression directed by nuclear siRNAs in plants.
[Part (a) data courtesy of Seth Darst; see N. Korzheva et al., 2000, Science289:619–625. Part (b) data from P. Cramer et al., 2001, Science292:1863, PDB ID 1i50. Part (c) data from K. J. Armache et al., 2003, P. Natl. Acad. Sci. USA100:6964, and D. A. Bushnell and R. D. Kornberg, 2003, P. Natl. Acad. Sci. USA100:6969.]
FIGURE 9-12Comparison of three-dimensional structures of bacterial and eukaryotic RNA polymerases. (a, b) These space-filling models are based on x-ray crystallographic analysis. (a) RNA polymerase from the bacterium T. aquaticus. The five subunits of the bacterial enzyme are distinguished by color. Only the N-terminal domains of the α subunits are included in this model. (b) Core RNA polymerase II from S. cerevisiae. Ten of the 12 subunits constituting yeast RNA polymerase II are shown in this model. Subunits that are similar in conformation to those in the bacterial enzyme are shown in the same colors. The C-terminal domain of the large subunit RPB1 was not observed in the crystal structure, but it is known to extend from the position marked with a red arrow. (RPB is the abbreviation for “RNA polymerase B,” which is an alternative way of referring to RNA polymerase II.) DNA entering the polymerases as they transcribe to the right is diagrammed. (c) Space-filling model of yeast RNA polymerase II including subunits 4 and 7. These subunits extend from the core portion of the enzyme shown in (b) near the region of the C-terminal domain of the large subunit.
[Part (a) data courtesy of Seth Darst; see N. Korzheva et al., 2000, Science289:619–625. Part (b) data from P. Cramer et al., 2001, Science292:1863, PDB ID 1i50. Part (c) data from K. J. Armache et al., 2003, P. Natl. Acad. Sci. USA100:6964, and D. A. Bushnell and R. D. Kornberg, 2003, P. Natl. Acad. Sci. USA100:6969.]
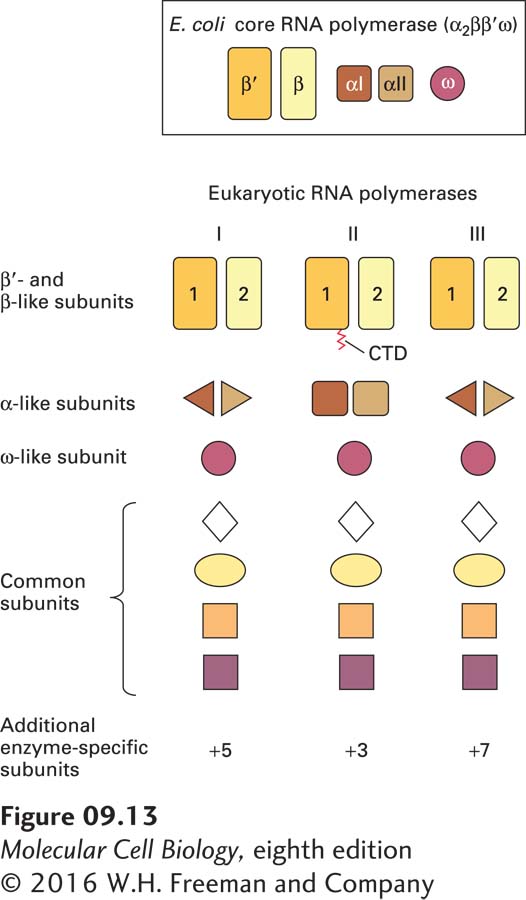
The two large subunits of all three eukaryotic RNA polymerases (and RNA polymerases IV and V of plants) are related to one another and are similar to the E. coli β′ and β subunits, respectively (see Figure 9-12a, b). Each of the eukaryotic RNA polymerases also contains an ω-like and two nonidentical α-like subunits (Figure 9-13). The extensive similarity in the structures of these core subunits in RNA polymerases from various sources indicates that RNA polymerase arose early in evolution and was largely conserved. This seems logical for an enzyme catalyzing a process as fundamental as the copying of RNA from DNA. In addition to the core subunits that are related to the E. coli RNA polymerase subunits, all three yeast RNA polymerases contain four additional small subunits, common to them but not to the bacterial RNA polymerase. Finally, each eukaryotic nuclear RNA polymerase has several enzyme-specific subunits that are not present in the other two (see Figure 9-13). Three of these additional subunits of Pol I and Pol III are homologous to the three additional Pol II-specific subunits. The other two Pol I-specific subunits are homologous to the Pol II general transcription factor TFIIF, discussed later, and the four additional subunits of Pol III are homologous to the Pol II general transcription factors TFIIF and TFIIE. These are likely stably associated with Pol III in the cell, and do not dissociate from it during purification.
Page 369
FIGURE 9-13Schematic representation of the subunit structure of the E. coli RNA core polymerase and yeast nuclear RNA polymerases. All three yeast polymerases have five core subunits homologous to the β, β′, two α, and ω subunits of E. coli RNA polymerase. The largest subunit (RPB1) of RNA polymerase II also contains an essential C-terminal domain (CTD). RNA polymerases I and III contain the same two nonidentical α-like subunits, whereas RNA polymerase II contains two other nonidentical α-like subunits. All three polymerases share the same ω-like subunit and four other common subunits. In addition, each yeast polymerase contains three to seven unique smaller subunits.
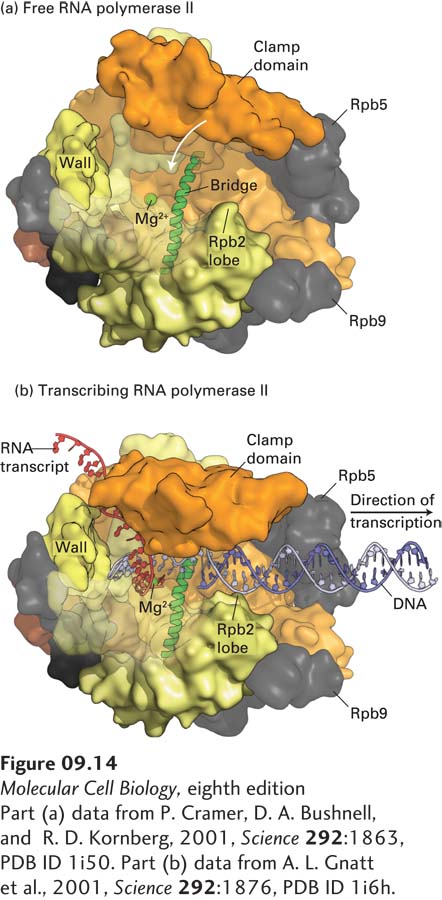
The clamp domain of subunit RPB1 is so designated because it has been observed in two different positions in crystals of free Pol II (Figure 9-14a) and in a complex that mimics the elongating form of the enzyme (Figure 9-14b). This domain rotates on a hinge that is probably open when downstream DNA is inserted into this region of the polymerase, and then swings shut when the enzyme is in its elongation mode. It is postulated that when the 8–9-bp RNA-DNA hybrid region near the active site (where RNA is base-paired to the template strand; see Figure 9-14b) is bound between RBP1 and RBP2, the clamp is locked in its closed position, anchoring the polymerase to the downstream double-stranded DNA. Furthermore, a transcription elongation factor called DSIF, discussed later, associates with the elongating polymerase, holding the clamp in its closed conformation. As a consequence, the polymerase is extraordinarily processive, which is to say that it continues to polymerize ribonucleotides until transcription is terminated. After termination and release of RNA from the exit channel, the clamp can swing open, releasing the enzyme from the template DNA. This mechanism can explain how human RNA polymerase II can transcribe the longest human gene, encoding dystrophin (DMD; see Figure 1-31), which is some 2 million base pairs in length, without dissociating and terminating transcription. Since transcription elongation proceeds at 1–2 kb per minute, transcription of the DMD gene requires approximately one day!
[Part (a) data from P. Cramer, D. A. Bushnell, and R. D. Kornberg, 2001, Science292:1863, PDB ID 1i50. Part (b) data from A. L. Gnatt et al., 2001, Science292:1876, PDB ID 1i6h.]
FIGURE 9-14The clamp domain of RPBI. The structures of the free (a) and transcribing (b) RNA polymerase II differ mainly in the position of a clamp domain in the RPB1 subunit (orange), which swings over the cleft between the jaws of the polymerase during formation of the transcribing complex, trapping the template DNA strand and transcript. Binding of the clamp domain to the 8–9-bp RNA-DNA hybrid may help couple clamp closure to the presence of RNA, stabilizing the closed, elongating complex. RNA is shown in red, and the template strand in light purple. For clarity, downstream nontemplate DNA is not shown. The clamp closes over the incoming downstream DNA. Portions of RBP2 that form one side of the cleft have been removed so that the nucleic acids can be better visualized. The Mg2+ ion that participates in catalysis of phosphodiester bond formation is shown in green. Wall is the domain of RPB2 that forces the template DNA entering the jaws of the polymerase to bend before it exits the polymerase. The bridge α helix, shown in green, extends across the cleft in the polymerase (see Figure 9-12b) and is postulated to bend and straighten as the polymerase translocates one base down the template strand. The nontemplate strand is thought to form a flexible single-stranded region above the cleft (not shown), extending from three bases downstream of the template base-paired to the 3′ base of the growing RNA to where the template strand exits the polymerase, where it hybridizes with the template strand to generate the transcription bubble.
[Part (a) data from P. Cramer, D. A. Bushnell, and R. D. Kornberg, 2001, Science292:1863, PDB ID 1i50. Part (b) data from A. L. Gnatt et al., 2001, Science292:1876, PDB ID 1i6h.]
Page 370
Gene-knockout experiments in yeast indicate that most of the subunits of the three nuclear RNA polymerases are essential for cell viability. Disruption of the genes encoding the few polymerase subunits that are not essential for viability (e.g., subunits 4 and 7 of RNA polymerase II) nevertheless results in very poorly growing cells. Thus all the subunits are necessary for eukaryotic RNA polymerases to function normally. Archaea, like eubacteria, have a single type of RNA polymerase involved in gene transcription, but archaeal RNA polymerases, like eukaryotic nuclear RNA polymerases, have on the order of a dozen subunits. Archaea also have general transcription factors, discussed later, that are related to those of eukaryotes, consistent with the closer evolutionary relationship between archaea and eukaryotes than between eubacteria and eukaryotes (see Figure 1-1).