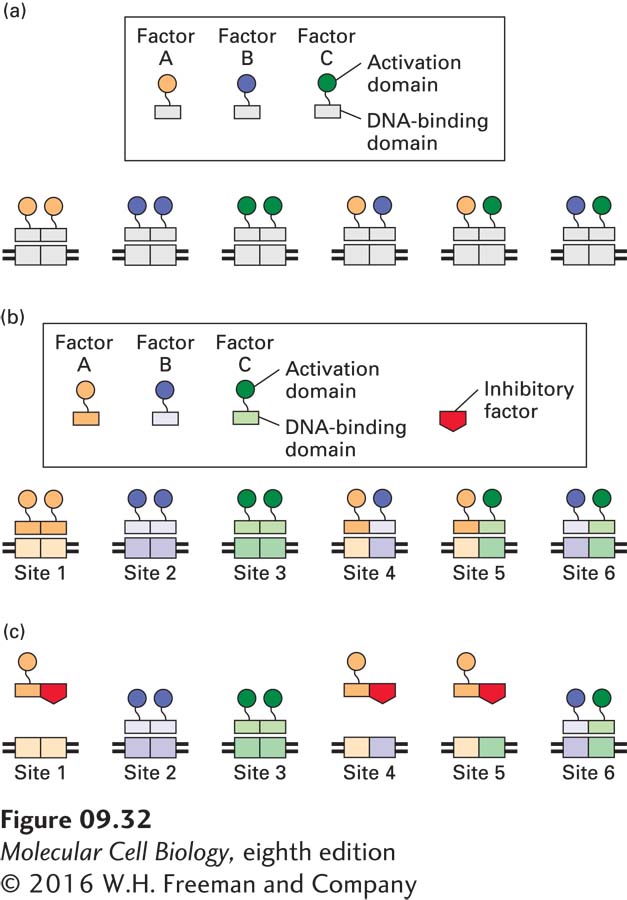
Two types of DNA-binding proteins discussed previously—bZIP and bHLH proteins—often exist in alternative heterodimeric combinations of monomers. Other classes of transcription factors not discussed here also form heterodimeric proteins. In some heterodimeric transcription factors, each monomer recognizes the same sequence. In these cases, the formation of alternative heterodimers does not increase the number of different sites on which the monomers can act, but rather allows the activation domains associated with each monomer to be brought together in alternative combinations that bind to the same site (Figure 9-32a). As we will see later, and in subsequent chapters, the activities of individual transcription factors can be regulated by multiple mechanisms. Consequently, a single bZIP- or bHLH-binding DNA regulatory element in the transcription-control region of a gene may elicit different transcriptional responses depending on which bZIP or bHLH monomers are expressed in the cell and how their activities are regulated.
FIGURE 9-32Combinatorial possibilities due to formation of heterodimeric transcription factors. (a) In some heterodimeric transcription factors, the activation domain of each monomer recognizes the same DNA sequence. In the hypothetical example shown, transcription-factor monomers A, B, and C can all interact with one another, creating six different alternative combinations of activation domains that can all bind at the same site. Each composite binding site is divided into two half-sites, and each heterodimeric factor contains the activation domains of its two constituent monomers. (b) When transcription-factor monomers recognize different DNA sequences, six alternative combinations of the transcription-factor monomers A, B, and C, each with a unique pair of activation domains, can bind to six different DNA sequences (sites 1–6). (c) Expression of an inhibitory factor (red) that interacts only with the dimerization domain of A inhibits binding; hence transcriptional activation at sites 1, 4, and 5 is inhibited, but activation at sites 2, 3, and 6 is unaffected.
In some heterodimeric transcription factors, however, each monomer has a different DNA-binding specificity. The resulting combinatorial possibilities increase the number of potential DNA sequences that a family of transcription factors can bind. Three different transcription-factor monomers could theoretically combine to form six different homo- and heterodimeric transcription factors, as illustrated in Figure 9-32b. Four different monomers could form a total of ten dimeric factors; five monomers, sixteen dimeric factors; and so forth. In addition, inhibitory factors are known that bind to some bZIP and bHLH monomers, thereby blocking their binding to DNA. When these inhibitory factors are expressed, they repress transcriptional activation by the factors with which they interact (Figure 9-32c). Thus the rules governing the interactions of members of a heterodimeric transcription factor family are complex. This combinatorial complexity expands both the number of DNA sites from which these factors can activate transcription and the ways in which they can be regulated.
Page 388
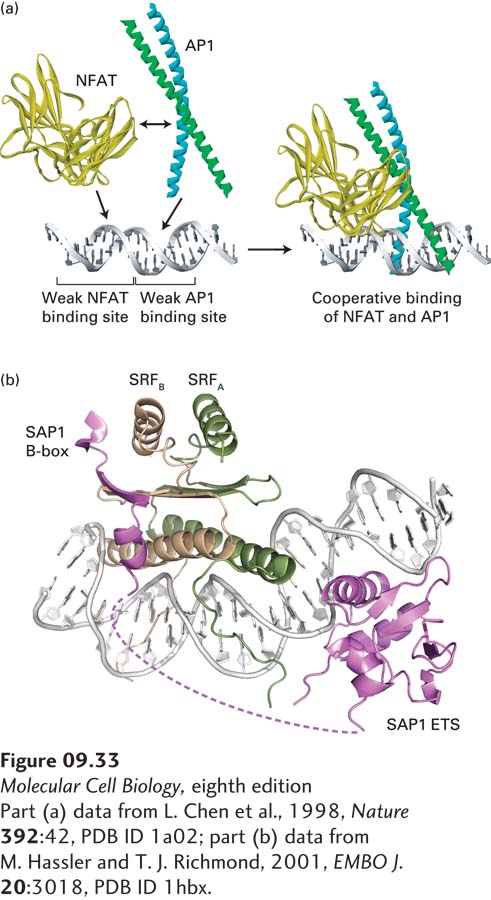
Similar combinatorial transcription regulation is achieved through the interaction of structurally unrelated transcription factors bound to closely spaced binding sites in DNA. An example is the interaction of two transcription factors, NFAT and AP1, that bind to neighboring sites in a composite promoter-proximal element regulating the gene encoding interleukin-2 (IL-2). Expression of the IL-2 gene is critical to the immune response, but abnormal expression of IL-2 can lead to autoimmune diseases such as rheumatoid arthritis (see Chapter 23). Neither NFAT nor AP1 binds to its site in the IL-2 control region in the absence of the other. The affinities of these factors for these particular DNA sequences are too low for the individual factors to form a stable complex with DNA. However, when both NFAT and AP1 are present, protein-protein interactions between them stabilize the ternary complex composed of NFAT, AP1, and DNA (Figure 9-33a). Such cooperative DNA binding by various transcription factors results in considerable combinatorial complexity of transcriptional control. As a result, the 1400 or so transcription factors encoded in the human genome can bind to DNA through a much larger number of cooperative interactions, resulting in unique transcriptional control for each of the roughly 21,000 human genes. In the case of IL-2, transcription occurs only when NFAT is activated, which results in its transport from the cytoplasm to the nucleus, and the two subunits of AP1 are synthesized. These two events are controlled by distinct signal transduction pathways (see Chapters 15 and 16), allowing stringent control of IL-2 expression.
[Part (a) data from L. Chen et al., 1998, Nature392:42, PDB ID 1a02; part (b) data from M. Hassler and T. J. Richmond, 2001, EMBO J.20:3018, PDB ID 1hbx.]
FIGURE 9-33Cooperative binding of two unrelated transcription factors to neighboring sites in a composite control element. (a) By themselves, both monomeric NFAT and heterodimeric AP1 transcription factors have low affinity for their respective binding sites in the IL-2 promoter-proximal region. Protein-protein interactions between NFAT and AP1 add to the overall stability of the NFAT-AP1-DNA complex, so that the two proteins bind to the composite site cooperatively. (b) Cooperative DNA binding by dimeric SRF and monomeric SAP1 can occur when their binding sites are separated by 5–30 bp and when the SAP1 binding site is inverted because the domain of SAP1 that interacts with SRF is connected to the DNA-binding domain of SAP1 by a flexible linker region of the SAP1 polypeptide chain (dotted line).
[Part (a) data from L. Chen et al., 1998, Nature392:42, PDB ID 1a02; part (b) data from M. Hassler and T. J. Richmond, 2001, EMBO J.20:3018, PDB ID 1hbx.]
Cooperative binding by NFAT and AP1 occurs only when their weak binding sites are positioned quite close to each other in DNA. The sites must be located at a precise distance from each other for effective binding. The requirements for cooperative binding are not so stringent in the case of some other transcription factors and transcription-control regions. For example, the EGR-1 control region contains a composite binding site to which the SRF and SAP1 transcription factors bind cooperatively (Figure 9-33b). Because SAP1 has a long, flexible domain that interacts with SRF, the two proteins can bind cooperatively when their individual sites in DNA are separated by any distance up to about 30 bp or are inverted relative to each other.