CONCEPT5.1 Biological Membranes Have a Common Structure and Are Fluid
The evolution of cellular life required the presence of a boundary, a way to separate the inside of the cell from the surrounding environment. This need was—and still is—fulfilled by biological membranes. Like many of the basic processes of life at the cellular level, the functions of membranes are carried out by a molecular structure shared by most organisms.
A biological membrane’s structure and functions are determined by the chemical properties of its constituents: lipids, proteins, and carbohydrates. An important concept that emerges from our consideration of these molecules is polarity and how polarity influences the way a molecule interacts with water. Recall from Concept 2.2 that some compounds are polar and hydrophilic (“water-loving”), whereas others are nonpolar and hydrophobic (“water-hating”), and that a phospholipid has both polar and nonpolar regions. The nonpolar regions of phospholipids and membrane proteins interact to form an insoluble barrier. The phospholipid bilayer serves as a lipid “lake” in which a variety of proteins “float” (FIGURE 5.1). This general design is known as the fluid mosaic model.
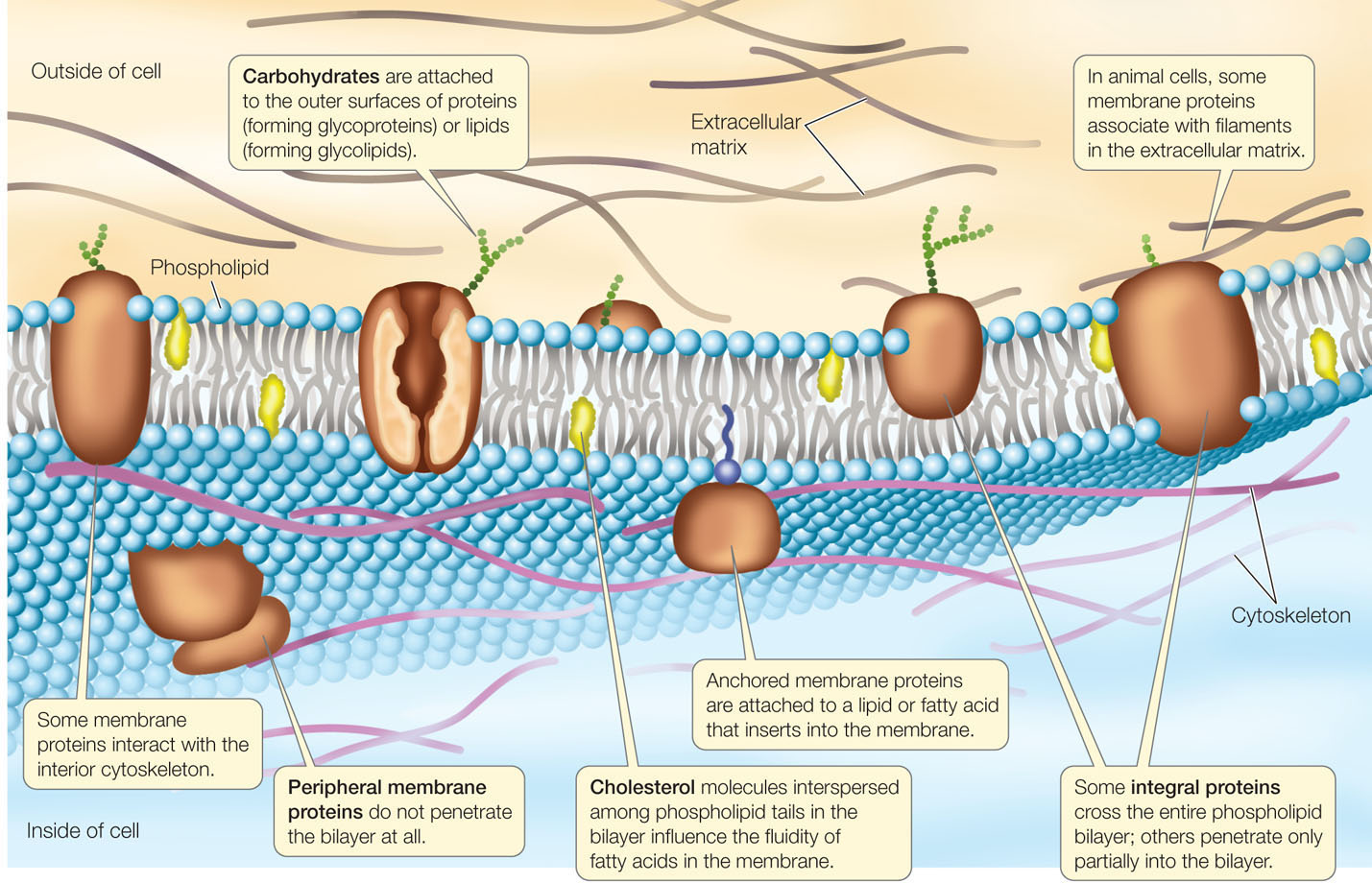
Membranes contain a wide array of proteins, most of which are noncovalently embedded in the phospholipid bilayer. These proteins are held within the membrane by their hydrophobic regions (also called their hydrophobic “domains”). The proteins’ hydrophilic regions are exposed to the watery conditions on one or both sides of the bilayer. Membrane proteins have three major functions: some move materials through the membrane, others are involved in intercellular recognition and adhesion, while others receive chemical signals from the cell’s external environment.
The carbohydrates associated with membranes are attached to either lipids or protein molecules. They are generally located on the outside of the cell, where they interact with substances in the external environment. Like some membrane proteins, carbohydrates are crucial for recognizing specific molecules, such as those on the surfaces of adjacent cells.
Each membrane has constituents that are suitable for the specialized functions of the cell or organelle it surrounds. As you read about the different molecules in membranes, keep in mind that some membranes contain many protein molecules, others are lipid-rich, others have significant amounts of cholesterol, and still others are rich in carbohydrates.
Lipids form the hydrophobic core of the membrane
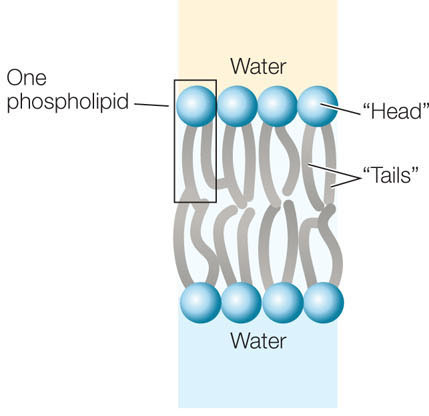
The lipids in biological membranes are usually phospholipids, with hydrophilic and hydrophobic regions:
- Hydrophilic regions. The phosphorus-containing “head” of a phospholipid is electrically charged and therefore associates with polar water molecules.
- Hydrophobic regions. The long, nonpolar fatty acid “tails” of a phospholipid associate with other nonpolar materials, but they do not dissolve in water or associate with hydrophilic substances.
One way in which phospholipids can coexist with water is to form a bilayer, with the fatty acid “tails” of the two layers interacting with each other, and the polar “heads” facing the outside, aqueous environment:
LINK
The properties of phospholipid bilayers are described in Concept 2.4
The thickness of a biological membrane is about 8 nm (0.008 μm), which is twice the length of a typical phospholipid. The page you are reading is about 3,000 times thicker than this.
As we noted in Chapter 4, it is possible in the laboratory to make artificial bilayers with the same organization as natural membranes. Small holes in such bilayers seal themselves spontaneously. The capacity of phospholipids to associate with one another and maintain a bilayer organization helps biological membranes fuse during vesicle formation, phagocytosis, and related processes (see Concept 4.3, especially Figure 4.9).
Although biological membranes all share a similar structure, there are many different kinds of phospholipids, and membranes from different cells or organelles may differ greatly in their lipid composition. Phospholipids can differ in terms of fatty acid chain length (number of carbon atoms), degree of unsaturation (number of double bonds) in the fatty acids, and the kinds of polar (phosphate-containing) groups present. The most common fatty acids in membranes have chains with 16–18 carbon atoms and 0–2 double bonds. Saturated fatty acid chains (those with no double bonds) allow close packing of phospholipids in the bilayer, whereas the “kinks” in the unsaturated fatty acids make for a less dense, more fluid packing (see Figure 2.12).
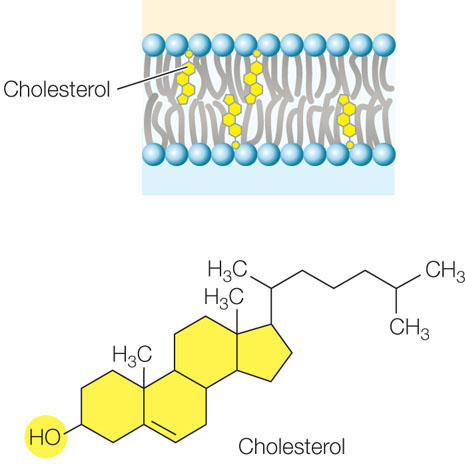
Up to 25 percent of the lipid content of an animal cell’s cell membrane may be the steroid cholesterol. Steroids are a family of carbon compounds that have multiple linked rings. Cholesterol plays an important role in modulating membrane fluidity (see below); other steroids function as hormones (see Concept 35.2). The hydroxyl group (—OH) on the cholesterol molecule interacts with the polar heads of the phospholipids, while the nonpolar rings insert among the fatty acid chains of the membrane:
The fatty acids of the phospholipids make the membrane somewhat fluid—about as fluid as olive oil. This fluidity permits some molecules to move laterally within the plane of the membrane. A given phospholipid molecule in the cell membrane can travel from one end of the cell to the other in a little more than 1 second!
Biological membranes have a common structure and are fluid
The membrane lipids of a cell can be labeled with a fluorescent tag so the entire surface of the cell will glow evenly under ultraviolet light. If a strong laser light is then shone on a tiny region of the cell, that region gets bleached (the strong light destroys the fluorescent tag) and there is a “hole” in the cell surface fluorescence (though not an actual hole in the cell’s membrane). After the laser is turned off, the hole gradually fills in with fluorescent lipids that diffuse in from other parts of the membrane. The time it takes for the “hole” to disappear is a measure of membrane fluidity. The table shows some data for cells with altered membrane compositions.a Explain the effect of each alteration.
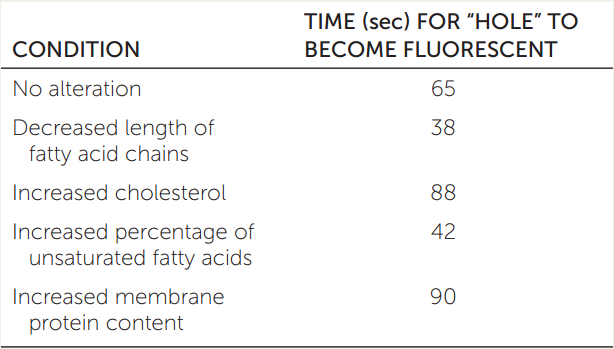
a Adapted from E. Wu et al. 1977. Biochemistry 16: 3936–3941.
85
Membrane fluidity is affected by several factors, two of which are particularly important:
- Lipid composition. Cholesterol and long-chain, saturated fatty acids pack tightly together, resulting in less fluid membranes. Unsaturated fatty acids or those with shorter chains tend to increase membrane fluidity. Some anesthetics are nonpolar and act by inserting into cell membranes. They reduce the fluidity of nerve cell membranes and thereby decrease nerve activity.
- Temperature. Membrane fluidity declines under cold conditions because molecules move more slowly at lower temperatures. For example, when your fingers get numb after contact with ice, it is due to a reduction in membrane fluidity in the nerve cells. To address this problem, some organisms simply change the lipid composition of their membranes when their environment gets cold, replacing saturated with unsaturated fatty acids and using fatty acids with shorter chains. These changes play a role in the survival of plants, bacteria, and hibernating animals during the winter.
While phospholipid molecules can easily move laterally within a membrane, it is rare for a phospholipid in one half of the bilayer to spontaneously flip over to the other side. For that to happen, the polar part of the molecule would have to move through the hydrophobic interior of the membrane. Since spontaneous flip-flops are rare, the inner and outer halves of the bilayer may be quite different in the kinds of phospholipids they contain.
Proteins are important components of membranes
All biological membranes contain proteins. Typically, cell membranes have about 1 protein molecule for every 25 phospholipid molecules. This ratio varies depending on membrane function. In the inner membrane of the mitochondrion, which is specialized for energy processing, there is 1 protein for every 5 lipids. By contrast, myelin—a membrane that encloses portions of some neurons (nerve cells) and acts as an electrical insulator—has only 1 protein for every 70 lipids.
Recall from Table 3.2 that some amino acids contain nonpolar, hydrophobic R groups, whereas others contain polar (or charged), hydrophilic R groups. The arrangement of these amino acids in a membrane protein determines whether the membrane protein will insert into the nonpolar lipid bilayer and how it will be positioned. There are two general types of membrane proteins:
- Peripheral membrane proteins lack exposed hydrophobic groups and are not embedded in the bilayer. Instead, they have polar or charged regions that interact with exposed parts of integral membrane proteins, or with the polar heads of phospholipid molecules (see Figure 5.1).
- Integral membrane proteins are at least partly embedded in the phospholipid bilayer. Like phospholipids, these proteins have both hydrophilic and hydrophobic regions:
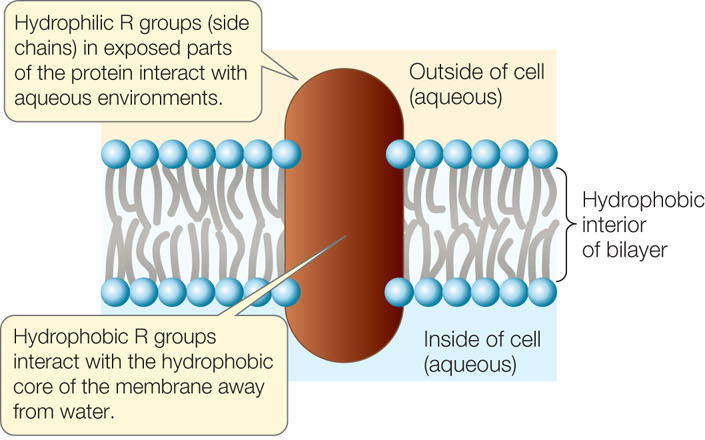
Membrane proteins and lipids generally interact only noncovalently. The polar ends of proteins can interact with the polar ends of lipids, and the nonpolar regions of both molecules can interact hydrophobically. However, some membrane proteins have fatty acids or other lipid groups covalently attached to them. These are referred to as anchored membrane proteins, because it is their hydrophobic lipid components that anchor them in the phospholipid bilayer (see Figure 5.1).
Proteins are asymmetrically distributed on the inner and outer surfaces of membranes. An integral membrane protein that extends all the way through the phospholipid bilayer and protrudes on both sides is known as a transmembrane protein. In addition to one or more transmembrane domains (regions) that extend through the bilayer, such a protein may have domains with other specific functions on the inner and outer sides of the membrane. Transmembrane proteins are always oriented the same way—domains with specific functions inside or outside the cell are always found on the correct side of the membrane. Peripheral membrane proteins are located on one side of the membrane or the other. This asymmetrical arrangement gives the two surfaces of the membrane different properties. As we will soon see, these differences have great functional significance.
Like lipids, some membrane proteins move relatively freely within the phospholipid bilayer. Cell fusion experiments illustrate this migration dramatically. When two cells fuse, a single continuous membrane forms around both cells, and some proteins from each cell distribute themselves uniformly around this membrane (FIGURE 5.2).
Investigation
HYPOTHESIS
Proteins embedded in a membrane can diffuse freely within the membrane.
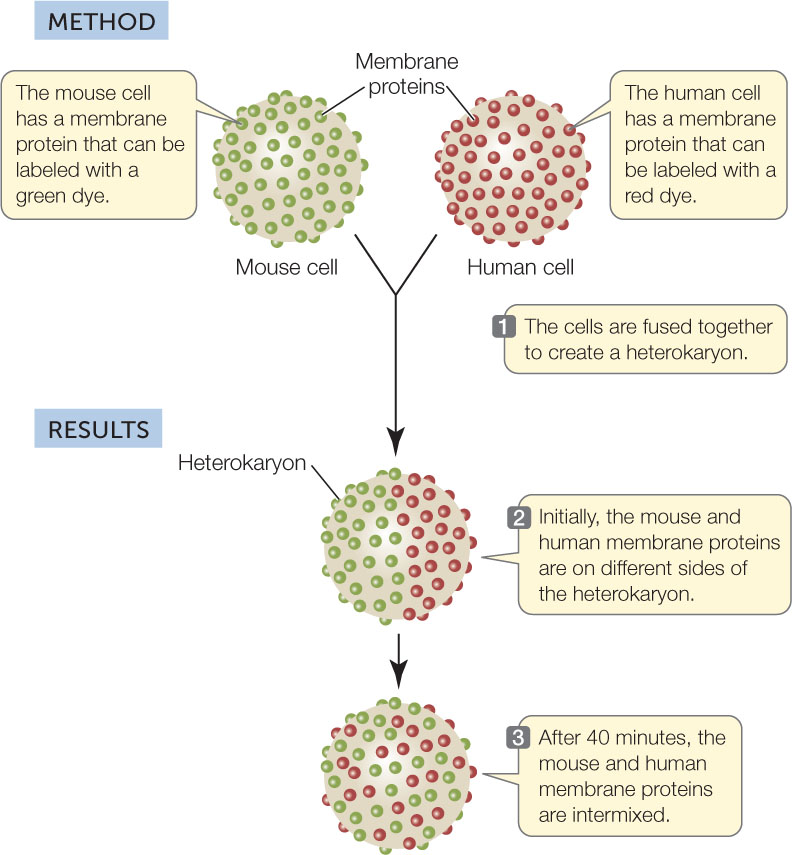
CONCLUSION
Membrane proteins can diffuse rapidly in the plane of the membrane.
ANALYZE THE DATA
The experiment was repeated at various temperatures with the following results:

Plot these data on a graph of Percentage Mixed vs. Temperature. Explain these data, relating the results to the concepts of diffusion and membrane fluidity.
a L. Frye and M. Edidin. 1970. Journal of Cell Science 7: 319-335.
Although some proteins are free to migrate throughout the membrane, others appear to be contained within specific regions. These membrane regions are like a corral of horses on a farm: the horses are free to move around within the fenced area but not outside it. For example, a muscle cell protein that recognizes a chemical signal from a neuron is normally found only within a specific region of the cell membrane, where the neuron meets the muscle cell.
How does this happen? Proteins inside the cell can restrict the movement of proteins within a membrane. Components of the cytoskeleton may be attached to membrane proteins protruding into the cytoplasm (see Figure 5.1). The stability of the cytoskeleton may thus restrict the movement of attached membrane proteins.
86
Cell membrane carbohydrates are recognition sites
In addition to lipids and proteins, the cell membrane contains carbohydrates. The carbohydrates are located on the outer surface of the cell membrane and may be covalently bonded to lipids or to proteins:
- A glycolipid consists of a carbohydrate covalently bonded to a lipid.
- A glycoprotein consists of one or more short carbohydrate chains covalently bonded to a protein. The bound carbohydrates are oligosaccharides, usually not exceeding 15 monosaccharide units in length (see Concept 2.3).
- A proteoglycan is a protein with even more carbohydrate molecules attached to it, and the carbohydrate chains are often longer than in glycoproteins.
These chains of monosaccharides can generate a large diversity of linear or branched structures. These diverse structures play roles in cell communication and cell adhesion. For example, the carbohydrates on some glycolipids change when cells become cancerous. This change may allow white blood cells to target cancer cells for destruction.
LINK
The activities of white blood cells are described in Chapter 39
Cells can stick together (adhere) due to interactions between similar carbohydrates on the outer surfaces of two cells, or between a carbohydrate on one cell and a membrane protein on another cell. Or, two proteins can interact directly:
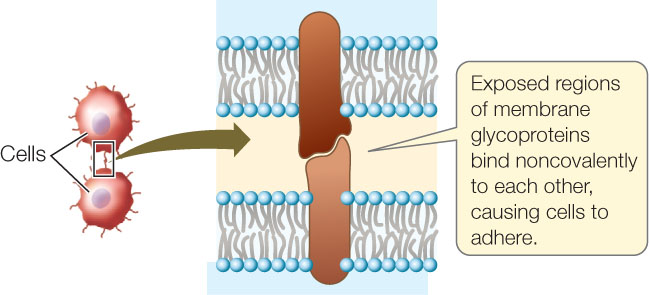
Cell adhesion occurs in all kinds of multicellular organisms.
Membranes are constantly changing
Membranes in eukaryotic cells are constantly forming, transforming from one type to another, fusing with one another, and breaking down. As we discussed in Concept 4.3, fragments of membrane move (in the form of vesicles) from the endoplasmic reticulum to the Golgi apparatus, and from the Golgi apparatus to the cell membrane (see Figure 4.8). Secondary lysosomes form when primary lysosomes from the Golgi apparatus fuse with phagosomes from the cell membrane (see Figure 4.9).
87
Because membranes interconvert so readily, we might expect all subcellular membranes to be chemically identical. However, that is not the case: there are major chemical differences among the membranes of even a single cell. Membranes are changed chemically when they form parts of certain organelles. In the Golgi apparatus, for example, the membranes of the cis face closely resemble those of the endoplasmic reticulum in chemical composition, but those of the trans face are more similar to the cell membrane.
CHECKpointCONCEPT5.1
- What are the differences between peripheral and integral membrane proteins?
- A membrane protein has the following amino acid sequence (see Table 3.2 for abbreviations):
EWDRHDFESGPTFIWLIWLVLAVLFLLLWAVLRPGKYKDKHE
Considering the R groups on the amino acids, predict the region of the protein that will be embedded within the membrane. - What is the evidence for membrane fluidity?
- If the cells of certain sponges are separated, they reaggregate because of binding between their membrane-associated proteoglycans. What would happen if the same experiment were conducted with cells treated to remove cell surface carbohydrates?
Now that you understand the structure of biological membranes, let’s see how their components function. In the sections that follow, we will focus on the cell membrane. We’ll look at how the cell membrane regulates the passage of substances that enter or leave a cell. Bear in mind that these principles also apply to the membranes that surround organelles.