CONCEPT4.3 Eukaryotic Cells Have a Nucleus and Other Membrane-Bound Compartments
Like prokaryotic cells, eukaryotic cells have a cell membrane, cytoplasm, and ribosomes. But as we noted earlier in this chapter, eukaryotic cells also have organelles within the cytoplasm whose interiors are separated from the cytosol by membranes (FIGURE 4.7).
66
67
68
Compartmentalization is the key to eukaryotic cell function
Each type of organelle has a specific role in the cell. Some organelles have been characterized as factories that make specific products. Others are like power plants that take in energy in one form and convert it into a more useful form. In addition, eukaryotic cells have some structures that are analogous to those seen in prokaryotes. For example, they have a cytoskeleton composed of protein fibers, and outside the cell membrane, an extracellular matrix.
When animal and plant cells are examined using electron microscopy, they have many organelles and structures in common—the most obvious is the cell nucleus. But they also have some differences. For example, many plant cells have chloroplasts that are colored green by the pigment used in photosynthesis. Figure 4.7 shows diagrams of an animal and a plant cell, with electron micrographs of some of the subcellular structures.
Ribosomes are factories for protein synthesis
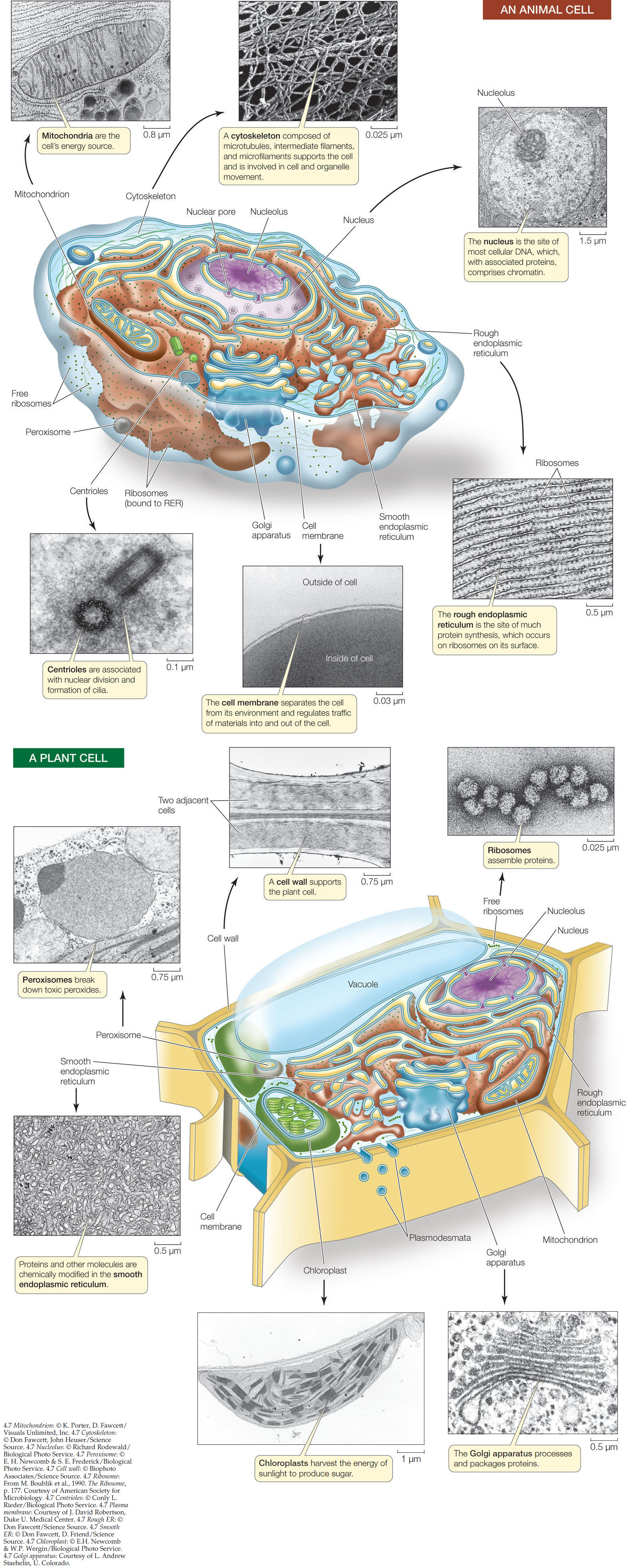
 Ribosomes translate the nucleotide sequence of a messenger RNA molecule into a polypeptide chain. The ribosomes of both prokaryotes and eukaryotes consist of one larger and one smaller subunit; the sizes of the subunits differ between the two cell types. Each subunit consists of one to three large RNA molecules called ribosomal RNA (rRNA) and multiple smaller protein molecules that are bound noncovalently to one another and to the rRNA. The ribosome is an amazingly precise structure. If the individual macromolecules are separated by disruption of their hydrophobic interactions, they will spontaneously reassemble into a functional complex.
Ribosomes translate the nucleotide sequence of a messenger RNA molecule into a polypeptide chain. The ribosomes of both prokaryotes and eukaryotes consist of one larger and one smaller subunit; the sizes of the subunits differ between the two cell types. Each subunit consists of one to three large RNA molecules called ribosomal RNA (rRNA) and multiple smaller protein molecules that are bound noncovalently to one another and to the rRNA. The ribosome is an amazingly precise structure. If the individual macromolecules are separated by disruption of their hydrophobic interactions, they will spontaneously reassemble into a functional complex.
LINK
Protein synthesis is described in more detail in Concept 10.4
Ribosomes are not membrane-enclosed compartments. In prokaryotic cells, ribosomes float freely in the cytoplasm. In eukaryotic cells they are found in the cytoplasm, where they may be free or attached to the surface of the endoplasmic reticulum (a membrane-enclosed organelle; see below), and also inside certain organelles—namely the mitochondria and the chloroplasts.
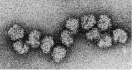
The nucleus contains most of the cell’s DNA
 As we noted in Chapter 3, hereditary information is stored in the sequence of nucleotides in DNA molecules. In eukaryotic cells, most of the DNA is in the nucleus. Most cells have a single nucleus, and it is usually the largest organelle; at 5 μm in diameter, the nucleus is substantially larger than most prokaryotic cells. The nucleus has several functions:
As we noted in Chapter 3, hereditary information is stored in the sequence of nucleotides in DNA molecules. In eukaryotic cells, most of the DNA is in the nucleus. Most cells have a single nucleus, and it is usually the largest organelle; at 5 μm in diameter, the nucleus is substantially larger than most prokaryotic cells. The nucleus has several functions:
- It is the location of the DNA and the site of DNA replication.
- It is where DNA is transcribed into RNA (see Concept 3.1).
- It contains the nucleolus, a region where ribosomes begin to assemble from RNA and proteins.
As you can see in Figure 4.7, the nucleus is enclosed by not one but two membranes: two lipid bilayers that together form the nuclear envelope. Functionally, this barrier separates DNA transcription (which occurs in the nucleus) from translation (which occurs in the cytoplasm). The two membranes of the nuclear envelope are perforated by thousands of nuclear pores, each measuring approximately 9 nm in diameter, which connect the interior of the nucleus to the cytoplasm. The pores regulate traffic between these two cellular compartments by allowing some molecules to enter or leave the nucleus and by blocking others. This allows the nucleus to regulate its information-processing functions.
Inside the nucleus, each DNA molecule is combined with proteins to form exceedingly long, thin threads called chromosomes. Different eukaryotic organisms have different numbers of chromosomes (ranging from two in one kind of Australian ant to hundreds in some plants). These DNA–protein complexes, which are also called chromatin, become much more compact during cell division, as you will see in Concept 7.2.
The outer membrane of the nuclear envelope folds outward into the cytoplasm and is continuous with the membrane of another organelle, the endoplasmic reticulum (see Figure 4.7).
The endomembrane system is a group of interrelated organelles
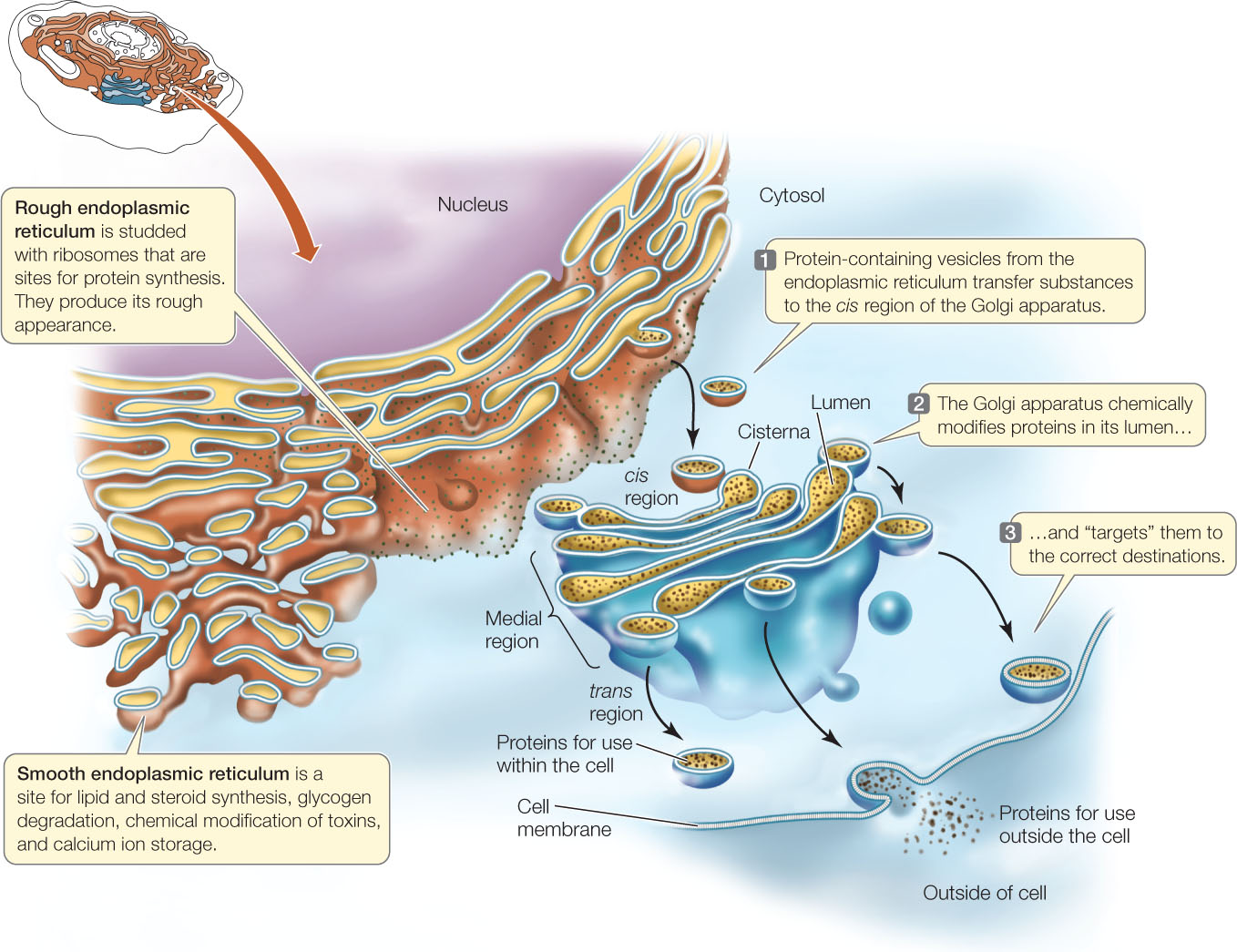
Much of the volume of many eukaryotic cells is taken up by an extensive endomembrane system. This interconnected system of membrane-enclosed compartments includes the nuclear envelope, endoplasmic reticulum, Golgi apparatus, and lysosomes, which are derived from the Golgi apparatus. Tiny, membrane-surrounded droplets called vesicles shuttle substances between the various components of the endomembrane system, as well as the cell membrane (FIGURE 4.8). In drawings and electron micrographs this system appears static, but in the living cell, membranes and the materials they contain are in constant motion. Membrane components have been observed to shift from one organelle to another within the endomembrane system. This suggests that all these membranes must be functionally related.
Endoplasmic Reticulum
Electron micrographs of eukaryotic cells reveal networks of interconnected membranes branching throughout the cytoplasm, forming tubes and flattened sacs about 1 μm across. These membranes are collectively called the endoplasmic reticulum, or ER. The interior compartment (lumen) of the ER is separate and distinct from the surrounding cytoplasm (see Figure 4.8). The ER can enclose up to 10 percent of the interior volume of the cell, and its extensive folding results in a surface area many times greater than that of the cell membrane. There are two types of ER: rough and smooth.
69
Eukaryotic cells have a nucleus and other membrane-bound compartments
Imagine that a group of scientists wanted to trace the path that the enzyme lipase follows between its site of synthesis and its final destination within a liver cell. First, the scientists exposed cultured liver cells to radioactive amino acids for 3 minutes. The amino acids entered the cells and became incorporated into all proteins synthesized during that time period. Then the radioactive amino acids were removed, so any proteins synthesized subsequently were not radioactive. At 5-minute intervals after the brief exposure to radioactive amino acids, some of the cells were broken open and fractionated to separate the organelles, as shown in Figure 4.4. An antibody that binds specifically to lipase was used to measure how much radioactive lipase was in each organelle at each time point (see Concept 39.4 for information on antibodies). The table shows the results.
- Plot percentage radioactive lipase versus time for each organelle. What can you conclude about the pathway of lipase in the cell after it is synthesized?
- Lipase breaks down lipids. Why is its organelle destination appropriate (see Figure 4.9)?
70
 The rough endoplasmic reticulum (RER) is called “rough” because of the many ribosomes attached to the outer surface of the membrane, giving it a rough appearance in electron micrographs. These ribosomes are not permanently attached to the ER but become attached when they begin synthesizing proteins destined for modification within the RER:
The rough endoplasmic reticulum (RER) is called “rough” because of the many ribosomes attached to the outer surface of the membrane, giving it a rough appearance in electron micrographs. These ribosomes are not permanently attached to the ER but become attached when they begin synthesizing proteins destined for modification within the RER:
- A protein enters the RER only if it contains a specific short sequence of amino acids that signals the ribosome to attach to the RER (see Concept 10.5).
- Once inside the RER, proteins are chemically modified to induce their three-dimensional functional shape and to chemically “tag” them for delivery to specific cellular destinations.
- The RER participates in transporting these proteins to other locations in the cell. The proteins are transported in vesicles that pinch off from the ER. All secreted proteins pass through the RER.
- Most membrane-bound proteins are made on the RER.
A polypetide that is synthesized on the RER surface is transported across the membrane and into the lumen while it is being translated. Once inside, it undergoes several changes, including the formation of disulfide bridges and folding into its tertiary structure. Many proteins are covalently linked to carbohydrate groups, thus becoming glycoproteins. These carbohydrate groups often have roles in recognition. For example, the carbohydrate groups on some secreted glycoproteins play roles in recognition and interactions between cells. Other carbohydrate groups “tag” their proteins for transfer to specific cellular locations. This “addressing” system is very important for ensuring that proteins arrive at their correct destinations. For example, the enzymes within the lysosomes (see below) are highly destructive and could destroy the cell if they were released into the cytosol.
 The smooth endoplasmic reticulum (SER) is connected to portions of the RER but lacks ribosomes and is more tubular (less like flattened sacs) than the RER. The SER has four important roles:
The smooth endoplasmic reticulum (SER) is connected to portions of the RER but lacks ribosomes and is more tubular (less like flattened sacs) than the RER. The SER has four important roles:
- It is responsible for the chemical modification of small molecules taken in by the cell that may be toxic to the cell. These modifications make the targeted molecules more polar, so they are more water-soluble and easily removed.
- It is the site for glycogen degradation in animal cells.
- It is the site where lipids and steroids are synthesized.
- It stores calcium ions, which when released trigger a number of cell responses, including muscle contraction.
LINK
The role of the SER in muscle contraction is described in Concept 33.1
Cells that synthesize a lot of protein for export are usually packed with RER. Examples include glandular cells that secrete digestive enzymes and white blood cells that secrete antibodies. In contrast, cells that carry out less protein synthesis (such as storage cells) contain less RER. Liver cells, which modify molecules (including toxins) that enter the body from the digestive system, have abundant SER.
Golgi Apparatus
 The Golgi apparatus is named after its discoverer, Camillo Golgi. It has two components: flattened membranous sacs called cisternae (singular cisterna), which are piled up like saucers in a stack about 1 μm thick, and small membrane-enclosed vesicles (see Figure 4.8). There can be many of these stacks in a cell.
The Golgi apparatus is named after its discoverer, Camillo Golgi. It has two components: flattened membranous sacs called cisternae (singular cisterna), which are piled up like saucers in a stack about 1 μm thick, and small membrane-enclosed vesicles (see Figure 4.8). There can be many of these stacks in a cell.
When protein-containing vesicles from the RER fuse with the Golgi apparatus membrane, the proteins are released into the lumen of a Golgi apparatus cisterna, where they may be further modified. The Golgi apparatus has several roles:
- It concentrates, packages, and sorts proteins before they are sent to their cellular or extracellular destinations.
- It adds some carbohydrates to proteins.
- It is where some polysaccharides for the plant cell wall are synthesized.
The cisternae of the Golgi apparatus have three functionally distinct regions: the cis region lies nearest to the nucleus or a patch of RER, the trans region lies closest to the cell membrane, and the medial region lies in between (see Figure 4.8). (The terms cis, trans. and medial derive from Latin words meaning “on the same side,” “on the opposite side,” and “in the middle,” respectively.) These three parts of the Golgi apparatus contain different enzymes and perform different functions.
Protein-containing vesicles from the ER fuse with the cis membrane of the Golgi apparatus. Other vesicles may transport proteins from one cisterna to the next, although it appears that some proteins move between cisterna through tiny channels. Vesicles budding off from the trans region carry their contents away from the Golgi apparatus. These vesicles go to the cell membrane or to the lysosome.
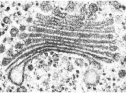
Lysosomes
The primary lysosomes originate from the Golgi apparatus. They contain hydrolases (digestive enzymes), and they are the sites where macromolecules—proteins, polysaccharides, nucleic acids, and lipids—are hydrolyzed into their monomers (see Chapter 2):
R1-R2 (linked monomers) + H2O → R1-OH + R2-H
For example,
Polypeptide + H2O → amino acids
A lysosome is about 0.5 μm in diameter; it is surrounded by a single membrane and has a densely staining, featureless interior (FIGURE 4.9). There may be dozens of lysosomes in a cell.
71
Some macromolecules that are hydrolyzed in lysosomes enter from the environment outside the cell by a process called phagocytosis (phago. “eat”; cytosis. “cellular”). In this process, a pocket forms in the cell membrane and then deepens and encloses material from outside the cell. The pocket becomes a small vesicle containing macromolecules (e.g., proteins), called a phagosome, which breaks free of the cell membrane to move into the cytoplasm. The phagosome fuses with a primary lysosome to form a secondary lysosome, in which hydrolysis occurs. The products of digestion (e.g., amino acids) pass through the membrane of the lysosome, providing monomers for other cellular processes. The “used” secondary lysosome, now containing undigested particles, then moves to the cell membrane, fuses with it, and releases the undigested contents to the environment.
Phagocytes are specialized cells whose major role is to take in and break down materials; they are found in nearly all animals and many protists. However, lysosomes are active even in cells that do not perform phagocytosis. All cells continually break down some of their components and replace them with new ones. The programmed destruction of cell components is called autophagy, and lysosomes are where the cell breaks down its own materials, even entire organelles, hydrolyzing their constituents.
How important is autophagy? An entire class of human diseases called lysosomal storage diseases occur when lysosomes fail to digest internal components; these diseases are often very harmful or fatal. For example, Tay-Sachs disease occurs when a particular lipid called a ganglioside is not broken down in the lysosomes and instead accumulates in brain cells and damages them. In the most common form of this disease, a baby starts exhibiting neurological symptoms and becomes blind, deaf, and unable to swallow after six months of age. Death occurs before age four.
Plant cells do not appear to contain lysosomes, but the central vacuole of a plant cell (which we will describe below) may function in an equivalent capacity because it, like lysosomes, contains many digestive enzymes.
Some organelles transform energy
A cell requires energy to make the molecules it needs for activities such as growth, reproduction, responsiveness, and movement. Mitochondria (found in all eukaryotic cells) harvest chemical energy, whereas chloroplasts (found in plants and other photosynthetic cells) harvest energy from sunlight.
Mitochondria
 In eukaryotic cells, the breakdown of energy-rich molecules such as the monosaccharide glucose begins in the cytosol. The molecules that result from this partial degradation enter the mitochondrion (plural mitochondria). whose primary function is to harvest the chemical energy of those molecules in a form the cell can use, namely the energy-rich nucleotide ATP (adenosine triphosphate). We will discuss these energy-harvesting processes in Chapter 6.
In eukaryotic cells, the breakdown of energy-rich molecules such as the monosaccharide glucose begins in the cytosol. The molecules that result from this partial degradation enter the mitochondrion (plural mitochondria). whose primary function is to harvest the chemical energy of those molecules in a form the cell can use, namely the energy-rich nucleotide ATP (adenosine triphosphate). We will discuss these energy-harvesting processes in Chapter 6.
72
A typical mitochondrion is somewhat less than 1.5 in diameter and 2–8 in length—about the size of many bacteria. It contains some DNA and can divide independently of the central nucleus. The number of mitochondria per cell ranges from one gigantic organelle in some unicellular protists to a few hundred thousand in large egg cells. An average human liver cell contains more than 1,000 mitochondria. Cells that are active in movement and growth require the most chemical energy, and these tend to have the most mitochondria per unit of volume.
Mitochondria have two membranes. The outer membrane has large pores, and most substances can pass through it. The inner membrane separates the biochemical processes of the mitochondrion from the surrounding cytosol. The inner membrane is extensively folded into structures called cristae, and the fluid-filled region inside the inner membrane is referred to as the mitochondrial matrix. The mitochondrion contains many enzymes for energy metabolism, as well as DNA and ribosomes for the synthesis of a small proportion of the mitochondrial proteins.
Plastids
 Plastids are present in the cells of plants and algae, and like mitochondria, they can divide autonomously. Plastids can differentiate into a variety of organelles, some of which are used for the storage of pigments (as in flowers), carbohydrates (as in potatoes), lipids, or proteins. An important type of plastid is the chloroplast, which contains the green pigment chlorophyll and is the site of photosynthesis (see Concepts 6.5 and 6.6). Photosynthesis is an anabolic process that converts light energy into the chemical energy contained in bonds between the atoms of carbohydrates.
Plastids are present in the cells of plants and algae, and like mitochondria, they can divide autonomously. Plastids can differentiate into a variety of organelles, some of which are used for the storage of pigments (as in flowers), carbohydrates (as in potatoes), lipids, or proteins. An important type of plastid is the chloroplast, which contains the green pigment chlorophyll and is the site of photosynthesis (see Concepts 6.5 and 6.6). Photosynthesis is an anabolic process that converts light energy into the chemical energy contained in bonds between the atoms of carbohydrates.
A chloroplast is enclosed within two membranes. In addition, it contains a series of internal membranes that look like stacks of flat, hollow discs, called thylakoids. Each stack of thylakoids is called a granum (plural grana). Light energy is converted to chemical energy on the thylakoid membranes. The aqueous fluid surrounding the thylakoids is called the stroma, and it is there that carbohydrates are synthesized. Like the mitochondrial matrix, the chloroplast stroma contains ribosomes and DNA, which are used to synthesize some of the chloroplast proteins.
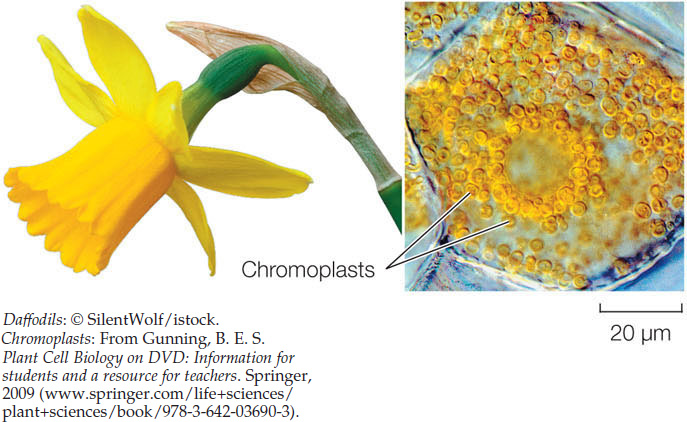
Other types of plastids have functions different from those of chloroplasts. Chromoplasts make and store red, yellow, and orange pigments, especially in flowers and fruits:
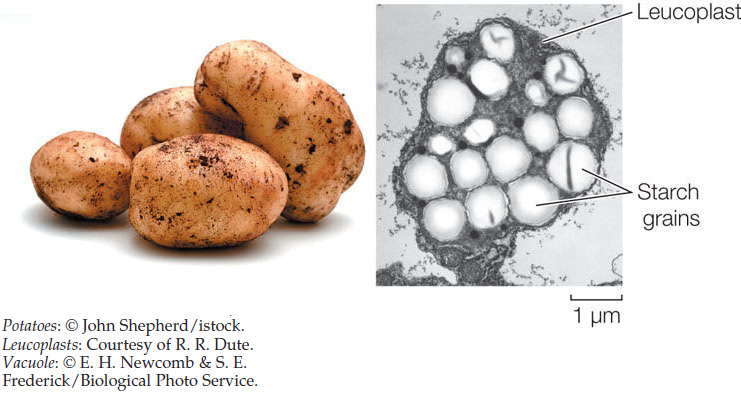
Leucoplasts are storage organelles for macromolecules such as starch:
Several other membrane-enclosed organelles perform specialized functions
There are several other kinds of membrane-bound organelles with specialized functions: peroxisomes, glyoxysomes, and vacuoles, including contractile vacuoles.
Peroxisomes
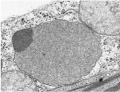 Peroxisomes are small (0.2–1 μm diameter) organelles that accumulate toxic peroxides, such as hydrogen peroxide (H2O2), which occur as the by-products of some biochemical reactions in all eukaryotes. These peroxides are safely broken down inside the peroxisomes without mixing with other components of the cell. A peroxisome has a single membrane and a granular interior containing specialized enzymes.
Peroxisomes are small (0.2–1 μm diameter) organelles that accumulate toxic peroxides, such as hydrogen peroxide (H2O2), which occur as the by-products of some biochemical reactions in all eukaryotes. These peroxides are safely broken down inside the peroxisomes without mixing with other components of the cell. A peroxisome has a single membrane and a granular interior containing specialized enzymes.
Glyoxysomes
Glyoxysomes are found only in plants. They are most abundant in young plants, which have many in each cell, and are the locations where stored lipids are converted into carbohydrates for transport to growing cells.
Vacuoles
 Vacuoles occur in many eukaryotic cells, but particularly those of plants and fungi (see Figure 4.7). There can be one large vacuole or many small ones in a cell. Plant vacuoles have several functions:
Vacuoles occur in many eukaryotic cells, but particularly those of plants and fungi (see Figure 4.7). There can be one large vacuole or many small ones in a cell. Plant vacuoles have several functions:
- Storage: Like all cells, plant cells produce a variety of toxic by-products and waste products. Plants store many of these in vacuoles. Because they are poisonous or distasteful, these stored materials deter some animals from eating the plants, and may thus contribute to the plants’ defenses and survival.
- Structure: In many plant cells, enormous vacuoles take up more than 90 percent of the cell volume and grow as the cell grows. The presence of dissolved substances in the vacuole causes water to enter it from the cytoplasm (which in turn takes up water from outside the cell), making the vacuole swell like a water-filled balloon. (This osmotic effect is illustrated in Figure 5.3.) The plant cell wall resists the swelling, causing the cell to stiffen from the increase in water pressure. This pressure is called turgor pressure, and it helps support the plant.
- Reproduction: Some pigments in the petals and fruits of flowering plants are contained in vacuoles. These pigments—the red, purple, and blue anthocyanins—are visual cues that help attract animals, which assist in pollination and seed dispersal.
- Catabolism: In the seeds of some plants, the vacuoles contain enzymes that hydrolyze stored seed proteins into monomers. The developing plant seedling uses these monomers as building blocks and sources of energy.
Many freshwater protists have contractile vacuoles. Their function is to get rid of the excess water that rushes into the cell because of the imbalance in solute concentration between the interior of the cell and its freshwater environment. The contractile vacuole enlarges as water enters, and then abruptly contracts, forcing the water out of the cell through a special pore structure.
CHECKpointCONCEPT4.3
- Make a table that summarizes eukaryotic cell organelles with regard to typical size, numbers per cell, and functions.
- What are some functions of the cell nucleus? What are the advantages of confining these functions within the nucleus, separated from the cytoplasm?
- Compare the structural and functional differences between rough and smooth endoplasmic reticulum.
- In I-cell disease, an enzyme in the endomembrane system that normally adds phosphorylated sugar groups to proteins is lacking, and the proteins are not targeted to the lysosomes as they would be in normal cells. The “I” stands for inclusion bodies that appear in the cells. What do you think these inclusions are, and why do they accumulate?
So far, we have discussed numerous membrane-enclosed organelles. Now we’ll turn to a group of cytoplasmic structures that do not directly involve membranes.