CONCEPT8.2 Alleles and Genes Interact to Produce Phenotypes
Phenotypes do not always follow the simple patterns of inheritance shown by the pairs of alleles for seed color and seed shape in peas, described in Concept 8.1. Existing alleles are subject to change by mutation and can give rise to new alleles—in fact, a single gene can have many alleles. In addition, alleles do not always show simple dominant–recessive relationships. A single allele may have multiple phenotypic effects, and a single character may be controlled by multiple genes. The expression of a gene is generally affected by interactions with other genes and with the environment.
New alleles arise by mutation
Genes are subject to mutations, which are rare, stable, and inherited changes in the genetic material. In other words, an allele can mutate (change) to become a different allele (this can happen in a number of different ways, as will be detailed in Concept 9.3). For example, we can envision that at one time all pea plants made round seeds and had the seed shape allele R. At some point, a mutation in R resulted in a new allele, r (wrinkled seeds). If this mutation was present in a cell that underwent meiosis, some of the resulting gametes would carry the r allele, and some offspring of this pea plant would carry the r allele in all of their cells.
Geneticists usually define one allele of a gene as the wild type; this is the allele that is present in most individuals in nature (“the wild”). Other alleles of that gene are usually called mutant alleles, and they may produce different phenotypes. The wild-type and mutant alleles are inherited according to Mendelian laws. A gene with a wild-type allele that is present less than 99 percent of the time (the rest of the alleles being mutant) is said to be polymorphic (Greek poly, “many”; morph, “form”).
LINK
By producing phenotypic variety, mutations provide raw material for evolution. An allele may become more or less prevalent in a population, depending on its effect on the fitness of the individuals carrying it; see Concept 15.3
Mendel developed his theories by studying just two alleles of each gene. But often a gene has multiple alleles (although any diploid individual will carry only two of them). The alleles may show a hierarchy of dominance when present in heterozygous individuals. An example is coat color in rabbits, determined by multiple alleles of the C gene (FIGURE 8.9):
- C determines dark gray.
- cchd determines chinchilla, a lighter gray.
- ch determines Himalayan (point restricted).
- c determines albino, no pigment.

The hierarchy of dominance for these alleles is C > cchd > ch > c. Any rabbit with the C allele (paired with itself or another allele) is dark gray, and a cc rabbit is albino. Intermediate colors result from different allele combinations, as shown in Figure 8.9. As this example illustrates, multiple alleles can increase the number of possible phenotypes.
Dominance is not always complete
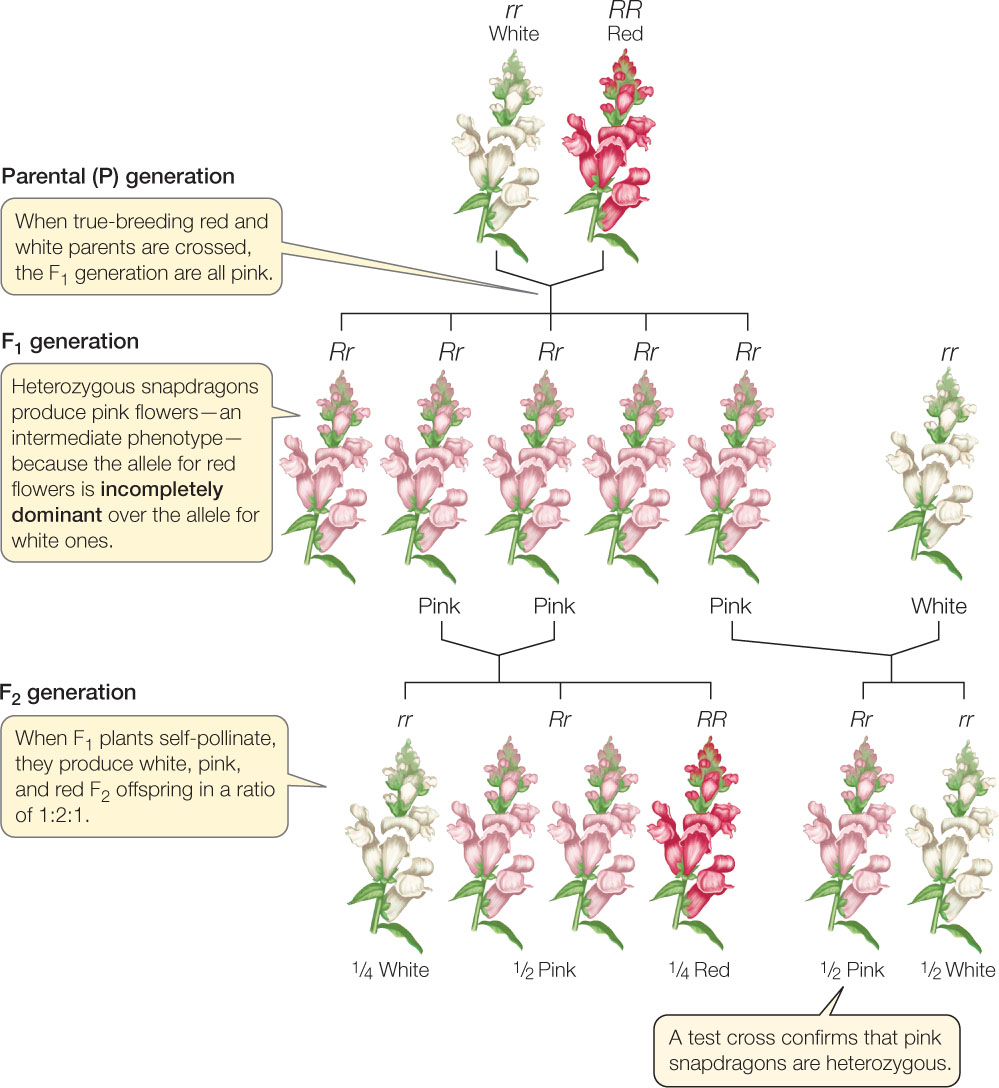
Many genes have alleles that are neither dominant nor recessive to one another. Instead, the heterozygotes have an intermediate phenotype, in a situation called incomplete dominance. For example, if a true-breeding red snapdragon is crossed with a white one, all the F1 flowers are an intermediate pink. Such cases appear to support the old blending theory of inheritance. However, further crosses indicate that this apparent blending can still be explained in terms of Mendelian genetics (FIGURE 8.10). The red and white snapdragon alleles have not disappeared, as those colors reappear in the F2 generation.
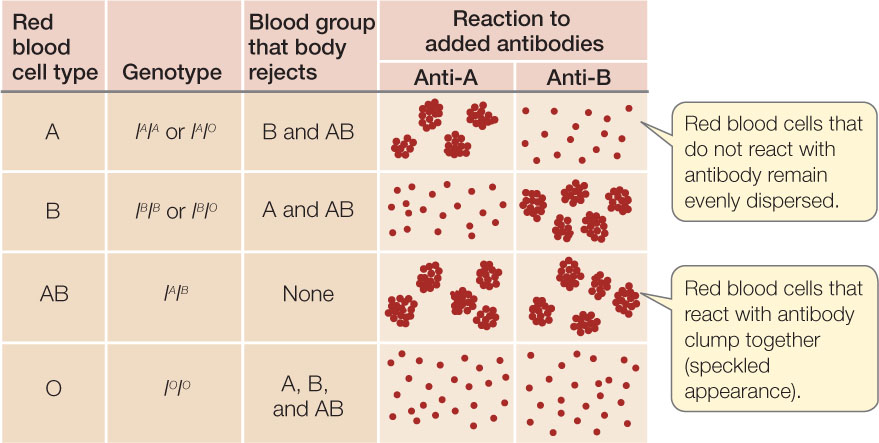
Sometimes two alleles of a gene both produce their phenotypes when present in a heterozygote—a phenomenon called codominance. An example is the ABO blood group in humans. The gene I encodes an enzyme involved in the attachment of sugars to a glycoprotein present on the surfaces of red blood cells. There are three alleles of the gene: IA, IB, and IO. The IA and IB alleles both encode active enzymes, but the enzymes attach different sugars to the glycoprotein. The IO allele does not encode an active enzyme, so no sugar is attached at that position on the glycoprotein. When two different alleles (e.g., IA and IB) are present, both alleles are expressed (both enzymes are made, so both types of glycoproteins are made). The A and B glycoproteins are antigenic: if a red blood cell with the A glycoprotein on its surface gets into the bloodstream of a person who lacks the IA allele, the recipient produces antibodies against the “nonself” cells (FIGURE 8.11). While the A and B glycoproteins are antigenic in people who do not have the IA or IB alleles, respectively, the O glycoprotein does not provoke antibody production. This makes people who are IOIO good blood donors.
160
LINK
Reactions against A or B glycoproteins are an adaptive immune response, as described in Concept 39.3
Genes interact when they are expressed
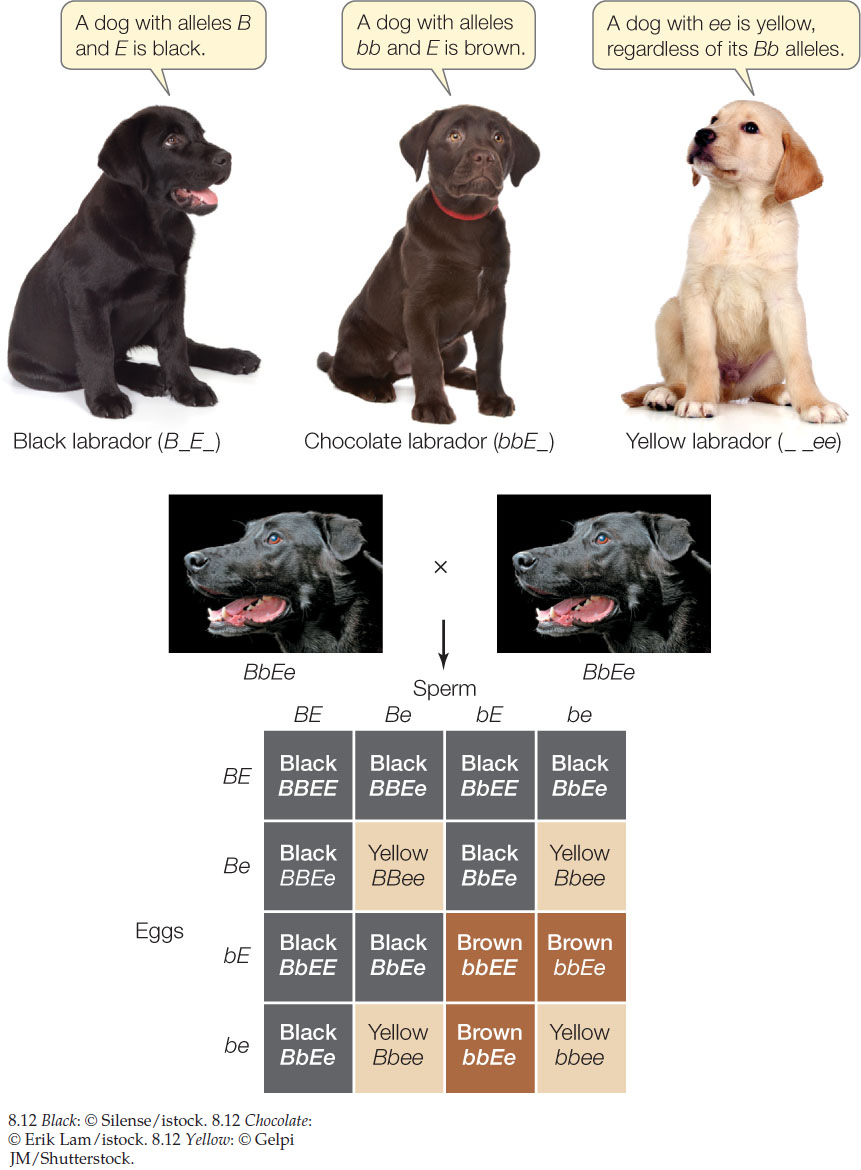
Epistasis occurs when the phenotypic expression of one gene is affected by another gene. For example, two genes (B and E) determine coat color in Labrador retrievers:
- Allele B (black pigment) is dominant to b (brown).
- Allele E (pigment deposition in hair) is dominant to e (no deposition, so hair is yellow).
An EE or Ee dog with BB or Bb is black; one with bb is brown. A dog with ee is yellow regardless of whether B or b alleles are present. Clearly, gene E determines the phenotypic expression of gene B, and is therefore epistatic to B (FIGURE 8.12).
Perhaps the most dramatic example of interacting genes or alleles is hybrid vigor (or heterosis). In 1876, Charles Darwin reported that when he crossed two different genetic varieties of corn, the offspring were 25 percent taller than either of the parent strains. Darwin’s observation was largely ignored for the next 30 years. In 1908, George Shull “rediscovered” this idea, reporting that not just plant height but the weight of the corn grain produced was dramatically higher in the offspring:

Agricultural scientists took note, and Shull’s paper had a lasting impact on the field of applied genetics. The cultivation of hybrid corn spread rapidly, and the practice of hybridization is now used for many other agricultural crops and animals. For example, beef cattle strains that are crossbred are larger and live longer than cattle bred within their own genetic strains.
What determines the “vigor” in hybrid vigor? A phenotype such as the amount of grain that a variety of corn produces in a given environment is determined by many genes and their alleles. Put more generally, most complex phenotypes are determined by multiple genes. Traits conferred by multiple genes are usually quantitative traits that need to be measured rather than assessed qualitatively. For example, grain yields must be measured, whereas a character such as Mendel’s pea seed color is an either-or quality that can be assessed by eye. The genetic basis of hybrid vigor is not well understood, but presumably the combinations of alleles and their products from two different varieties can interact to produce more vigorous offspring.
The environment affects gene action
The phenotype of an individual does not always result from its genotype alone. Genotype and environment often interact to determine the phenotype of an organism. It is especially important to remember this fact in the era of genome sequencing (see Chapter 12). When the sequence of the human genome was completed in 2003, it was hailed as the “book of life,” and public expectations of the benefits gained from this knowledge were (and are) high. But this kind of “genetic determinism”—the idea that an organism’s genome sequence determines all of its phenotype—is wrong. Common knowledge tells us that environmental variables such as light, temperature, and nutrition can affect the phenotypic expression of a genotype. For example, in humans body weight is determined not only by multiple genes but also by nutrition and activity.
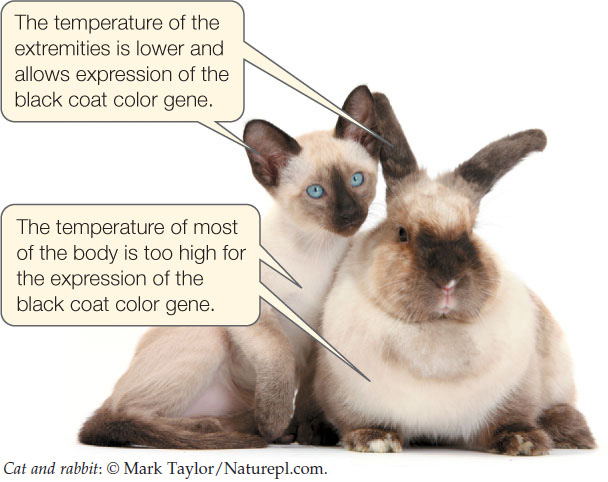
A common example of the interaction of genes and environment involves “point restriction” coat patterns found in Siamese cats and certain rabbit breeds. These animals carry the ch allele (see Figure 8.9), a mutant version of the C gene that controls coat color. As a result of this mutation, the enzyme encoded by the gene is inactive at temperatures above a certain point—usually 35°C. The animals maintain a body temperature above this, so their body fur is light. However, their extremities—feet, ears, nose, and tail—are cooler (around 25°C), and the fur on these regions is dark:
161
Alleles and genes interact to produce phenotypes
- In the genetic cross AaBbCcDdEE × AaBBCcDdEe where all the genes are on separate chromosomes, what fraction of the offspring will be heterozygous for all of these genes?
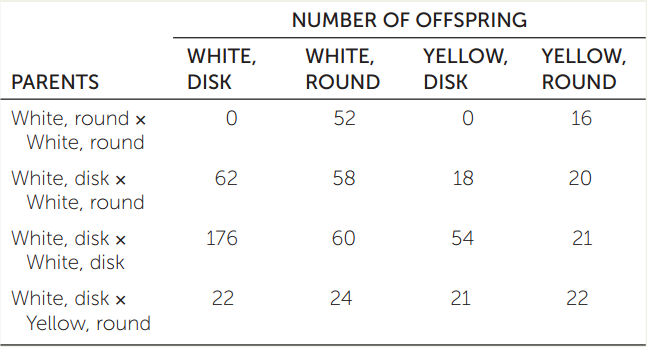
- In squash plants, fruit color is determined by one gene (W: white, dominant to w: yellow) and fruit shape by another gene on a different chromosome (D: disk shape, dominant to d: round). What were the genotypes of the parents for each cross in the table at right?
- In chickens, when the dominant alleles of the genes for rose comb (R) and pea comb (A) are present together, the bird has a walnut comb. Birds that are homozygous recessive for both genes (i.e., genotype ra) have a single comb. A rose-combed bird mated with a walnut-combed bird, and the phenotypes of the 8 offspring were:
⅜ walnut : ⅜ rose : ⅛ pea : ⅛ single.
What were the genotypes of the parents?
A simple experiment shows that the dark fur is temperature-dependent. If a patch of white fur on a point-restricted animal is shaved off and an ice pack is placed on the skin where the patch was, the fur that grows in is dark. This indicates that although the allele for dark fur was expressed all along, the environment inhibited the activity of the mutant enzyme. These animals are all white at birth because the extremities are kept warm in the mother’s womb.
162
Two parameters describe the effects of genes and environment on phenotype:
- Penetrance is the proportion of individuals in a group with a given genotype that actually show the expected phenotype. For example, many people who inherit a mutant allele of the gene BRCA1 develop breast cancer in their lifetimes. But for reasons that are not yet clear and must involve other genes and/or the environment, some people with the mutation do not develop breast cancer. So the BRCA1 mutation is said to be incompletely penetrant.
- Expressivity is the degree to which a genotype is expressed in an individual. For example, a woman with the BRCA1 allele may develop both breast and ovarian cancer as part of the phenotype, but another woman with the same mutation may only get breast cancer. So the mutation is said to have variable expressivity.
In a population of an organism, there is often wide variability with regard to a particular phenotype. Consider, for example, height in humans:

Human height is a quantitative trait. Many genes contribute to height, and the interactions between these genes are complex. But the environment also contributes to variation in height; for example, some people have better nutrition than others, and this can affect their growth. The heritability of a character is the relative contribution of genetic versus environmental factors to the variation in that character in a particular population. Typically, heritability varies from 0 to 1. For example, the heritability of human height varies from about 0.65 to 0.8, depending on the population studied. In a population where heritability is 0.65, 65 percent of the variation in height is due to genetic factors, while the remaining 35 percent is due to environmental effects. It is important to note that heritability estimates apply to variations within populations. They cannot be used to estimate the contribution of genetics to particular characters in an individual.
Heritability estimates are important for breeders of plants and animals because they provide information about whether it is more worthwhile to modify the environment or do genetic crosses to improve a phenotype.
CHECKpointCONCEPT8.2
- What is the difference between incomplete dominance and codominance?
- A point-restricted rabbit (see Figure 8.9) was mated with a chinchilla rabbit. The two offspring were albino and chinchilla. What were the genotypes of the parents?
- If a dominant allele of one gene, A, is necessary for hearing in humans, and the dominant allele of another gene, B, results in deafness regardless of the presence of other genes, what fraction of offspring in a pairing of AaBb and Aabb individuals will be deaf?
- Give an example from your own experience of a genotype whose expression is affected by the environment.
So far we have considered genes that obey Mendel’s law of independent assortment. But many genes are inherited together, with multiple genes transmitted as one unit. This apparent anomaly can be explained by the presence of multiple genes on a single chromosome.