CONCEPT17.4 Reproductive Isolation Is Reinforced When Diverging Species Come into Contact
As discussed in Concept 17.2, once a barrier to gene flow is established, reproductive isolation will begin to develop through genetic divergence. Over many generations, differences accumulate in the isolated lineages, reducing the probability that individuals from each lineage will mate successfully with one another when they come back into contact. In this way, reproductive isolation can evolve as a by-product of the genetic changes in the two diverging lineages.
Reproductive isolation may be incomplete when the incipient species come back into contact, however, in which case some hybridization will occur. If hybrid individuals are less fit than non-hybrids, selection favors parents that do not produce hybrid offspring. Under these conditions, selection results in strengthening, or reinforcement, of isolating mechanisms that prevent hybridization.
Mechanisms that prevent hybridization from occurring are called prezygotic isolating mechanisms. Mechanisms that reduce the fitness of hybrid offspring are called postzygotic isolating mechanisms. Postzygotic mechanisms result in selection against hybridization, which leads to the reinforcement of the prezygotic mechanisms.
Prezygotic isolating mechanisms prevent hybridization between species
Prezygotic isolating mechanisms, which come into play before fertilization, can prevent hybridization in several ways.
Mechanical Isolation
Differences in the sizes and shapes of reproductive organs may prevent the union of gametes from different species. With animals, there may be a match between the shapes of the reproductive organs of males and females of the same species, so that reproduction between species with mismatched structures is not physically possible. In plants, mechanical isolation may involve a pollinator. For example, orchids of the genus Ophrys produce flowers that look and smell like the females of particular species of wasps (FIGURE 17.8). When a male wasp visits and attempts to mate with the flower (mistaking it for a female wasp of his species), his mating behavior results in the transfer of pollen to and from his body by appropriately configured anthers and stigmas on the flower. Insects that visit the flower but do not attempt to mate with it do not trigger the transfer of pollen between the insect and the flower.

Temporal Isolation
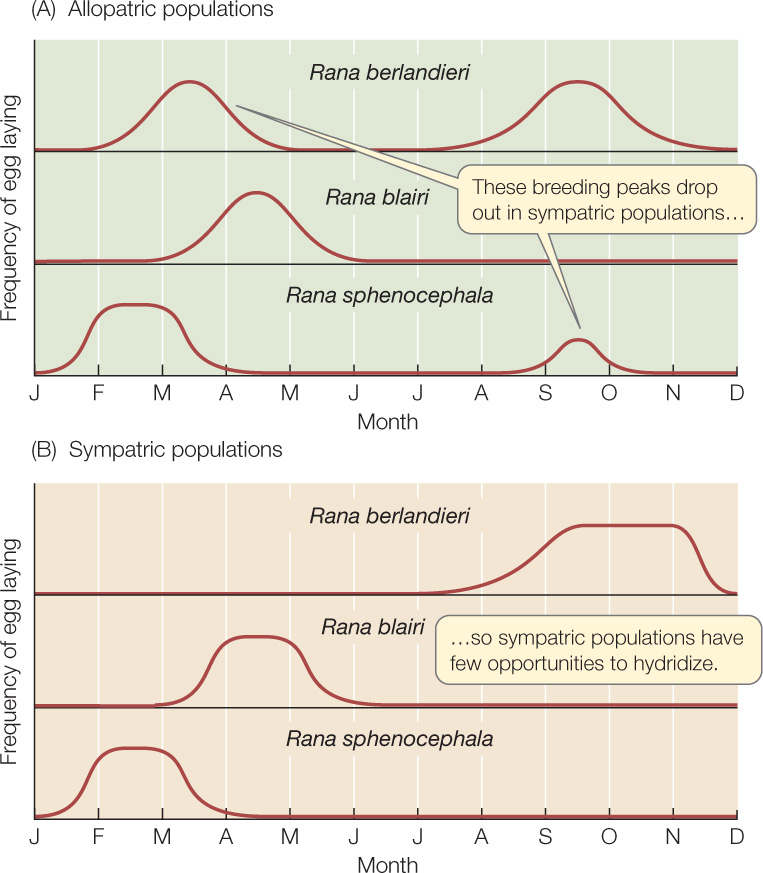
Many organisms have distinct mating seasons. If two closely related species breed at different times of the year (or different times of day), they may never have an opportunity to hybridize. For example, in sympatric populations of three closely related leopard frog species, each species breeds at a different time of year (FIGURE 17.9). Although there is some overlap in the breeding seasons, the opportunities for hybridization are minimal.
Behavioral Isolation
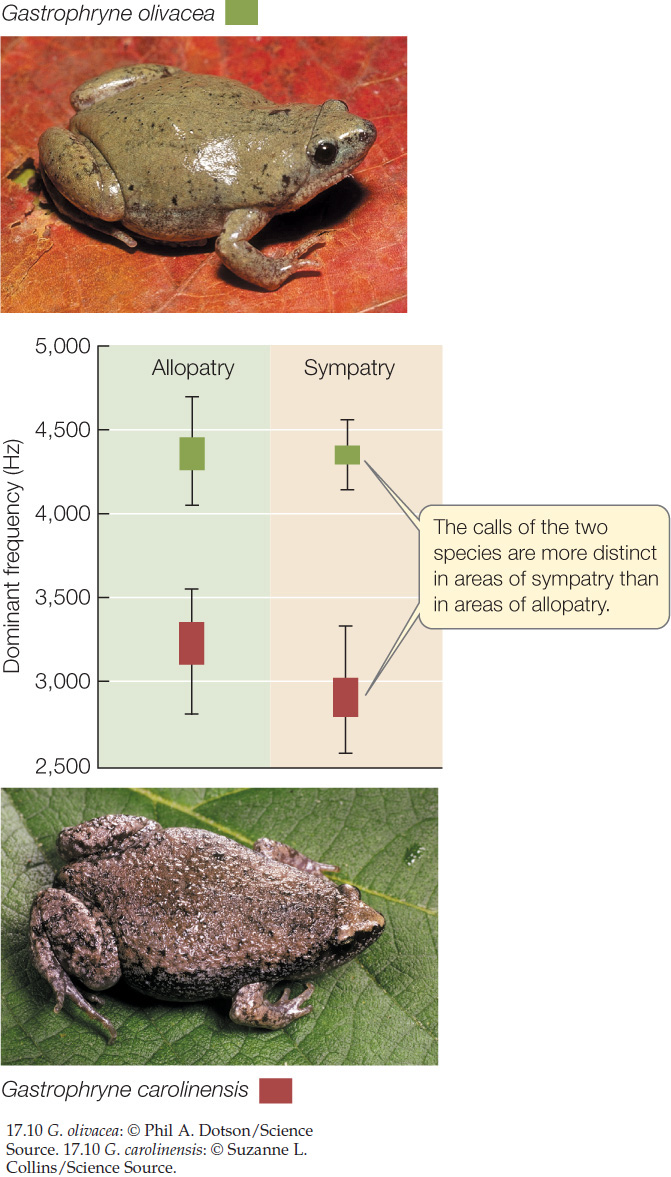
Individuals may reject, or fail to recognize, individuals of other species as potential mating partners. For example, the mating calls of male frogs of related species diverge quickly (FIGURE 17.10). Female frogs respond to mating calls from males of their own species but ignore the calls of other species, even closely related ones. The evolution of female preferences for certain male coloration patterns among the cichlids of Lake Malawi, described at the opening of this chapter, is another example of behavioral isolation.
352
Sometimes the mate choice of one species is mediated by the behavior of individuals of other species. For example, whether or not two plant species hybridize may depend on the food preferences of their pollinators. The floral traits of plants, including their color and shape, can enhance reproductive isolation either by influencing which pollinators are attracted to the flowers or by altering where pollen is deposited on the bodies of pollinators. A plant whose flowers are pendant (hanging downward; FIGURE 17.11A) will be pollinated by an animal with different physical characteristics than will a plant in which the flowers grow upright (FIGURE 17.11B). Because each pollinator prefers (and is adapted to) a different type of flower, the pollinators rarely transfer pollen from one plant species to the other.
Such isolation by pollinator behavior is seen in the mountains of California in two sympatric species of columbines (Aquilegia) that have diverged in flower color, structure, and orientation. Aquilegia formosa (FIGURE 17.11C) has pendant flowers with short spurs (spikelike, nectar-containing structures) and is pollinated by hummingbirds. A. pubescens (FIGURE 17.11D) has upright, lighter-colored flowers with long spurs and is pollinated by hawkmoths. The difference in pollinators means that these two species are effectively reproductively isolated even though they populate the same geographic range.

Habitat Isolation
When two closely related species evolve preferences for living or mating in different habitats, they may never come into contact during their respective mating periods. The Rhagoletis flies discussed in Concept 17.3 experienced such habitat isolation, as did the cichlid fishes that first adapted to rocky and sandy habitats upon entering Lake Malawi, as described at the opening of this chapter.
353
LINK
Some plants and their pollinators become so tightly adapted to each other that they develop mutually dependent relationships, as described in Concept 21.5
Gametic Isolation
The sperm of one species may not attach to the eggs of another species because the eggs do not release the appropriate attractive chemicals. Furthermore, the sperm may be unable to penetrate the egg because the two gametes are chemically incompatible. Thus even though the gametes of two species may come into contact, the gametes never fuse into a zygote.
Gametic isolation is extremely important for many aquatic species that spawn (release their gametes directly into the environment). For example, gametic isolation has been extensively studied in spawning sea urchins. A protein known as bindin is found in sea urchin sperm and functions in attaching (“binding”) the sperm to eggs. All sea urchin species studied produce this egg-recognition protein, but the bindin gene sequence diverges so rapidly that it becomes species-specific. Since sperm can only attach to eggs of the same species, no interspecific hybridization occurs.
Postzygotic isolating mechanisms result in selection against hybridization
Genetic differences that accumulate between two diverging lineages may reduce the survival and reproductive rates of hybrid offspring in any of several ways:
- Low hybrid zygote viability. Hybrid zygotes may fail to mature normally, either dying during development or developing phenotypic abnormalities that prevent them from becoming reproductively capable adults.
- Low hybrid adult viability. Hybrid offspring may have lower survivorship than non-hybrid offspring.
- Hybrid infertility. Hybrids may mature into infertile adults. For example, the offspring of matings between horses and donkeys—mules—are sterile. Although otherwise healthy, mules usually produce no descendants.
Natural selection does not directly favor the evolution of postzygotic isolating mechanisms. But if hybrids are less fit, individuals that breed only within their own species will leave more surviving offspring than will individuals that interbreed with another species. Therefore individuals that can avoid interbreeding with members of other species will have a selective advantage, and any trait that contributes to such avoidance will be favored.
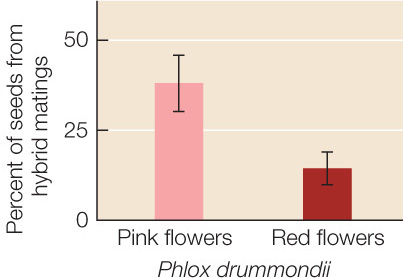
Donald Levin of the University of Texas has studied reinforcement of prezygotic isolating mechanisms in flowers of the genus Phlox. Levin noticed that individuals of Phlox drummondii in most of the range of the species in Texas usually have pink flowers. However, where P. drummondii is sympatric with its close relative the pink-flowered P. cuspidata, P. drummondii usually has red flowers. No other Phlox species has red flowers. Levin performed an experiment whose results showed that reinforcement may explain why red flowers are favored where the two species are sympatric (FIGURE 17.12).
354
Investigation
HYPOTHESIS
Red-flowered P. drummondii are less likely to hybridize with P. cuspidata than are pink-flowered P. drummondii.
- Introduce equal numbers of red-flowered and pink-flowered P. drummondii individuals into an area where many pink-flowered P. cuspidata are growing.

- After the flowering season ends, measure hybridization by assessing the genetic composition of the seeds produced by P. drummondii plants of both colors.
Of the seeds produced by pink-flowered P. drummondii, 38% were hybrids with P. cuspidata. Only 13% of the seeds produced by red-flowered P. drummondii were hybrids with P. cuspidata. Error bars show 95% confidence intervals (see Appendix B).
CONCLUSION
P. drummondii and P. cuspidata are less likely to hybridize if the flowers of the two species differ in color.
ANALYZE THE DATA
Data from Levin’s experiment show that the frequency of hybridization between Phlox drummondii and P. cuspidata was strongly dependent on the former’s flower color.

- The data reveal that red-flowered P. drummondii produced more seeds (208) than pink- flowered plants did (139), even though equal numbers of individuals of the two flower types were used. Does this difference influence your interpretation of Levin’s results? Why or why not?
- How would you improve the experimental design of this study? Should replicate or control sites be added? What kinds of additional test sites and conditions would you add?
aD. A. Levin. 1985. Evolution 39: 1275–1281.
Likely cases of reinforcement are often detected by comparing sympatric and allopatric populations of potentially hybridizing species, as in the case of Phlox. If reinforcement is occurring, then sympatric populations of closely related species are expected to evolve more effective prezygotic reproductive barriers than do allopatric populations of the same species. As Figure 17.9 shows, the breeding seasons of sympatric populations of different leopard frog species overlap much less than do those of the corresponding allopatric populations. Similarly, the frequencies of the frog mating calls illustrated in Figure 17.10 are more divergent in sympatric populations than in allopatric populations. In both cases, there appears to have been natural selection against hybridization in areas of sympatry.
Hybrid zones may form if reproductive isolation is incomplete
Unless reproductive isolation is complete, closely related species may hybridize in areas where their ranges overlap, resulting in the formation of a hybrid zone. When a hybrid zone first forms, most hybrids are offspring of crosses between purebred individuals of the two species. However, subsequent generations will include a variety of individuals with varying proportions of their genes derived from the original two species. Thus hybrid zones often contain recombinant individuals resulting from many generations of hybridization.
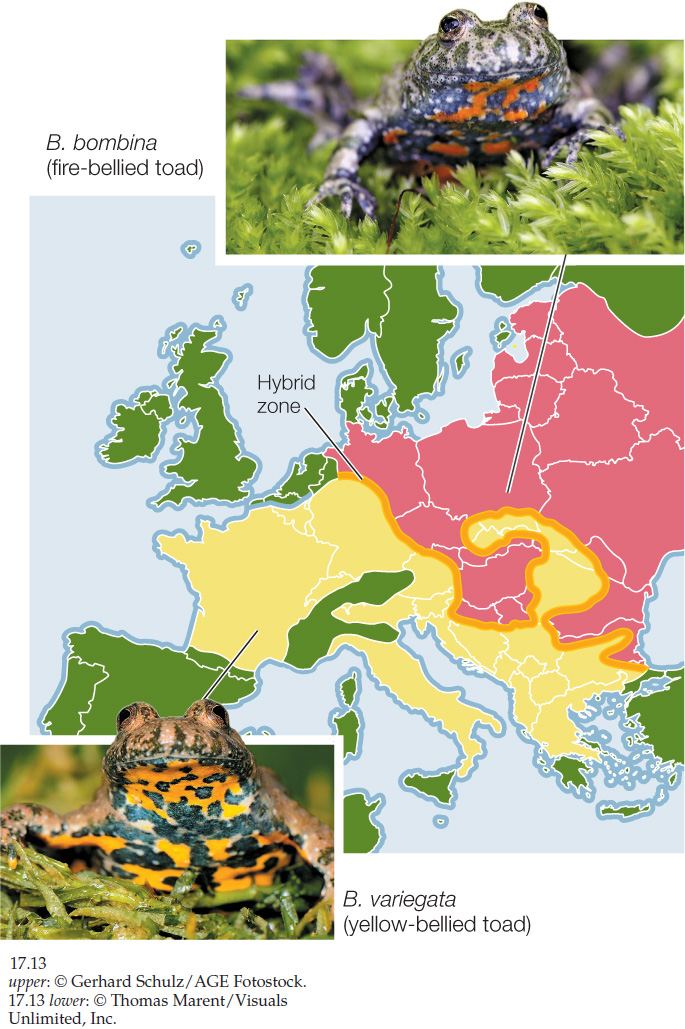
Detailed genetic studies can tell us much about why narrow hybrid zones may persist for long periods between the ranges of two species. In Europe, the hybrid zone between two toad species of the genus Bombina has been studied intensively. The fire-bellied toad (B. bombina) lives in eastern Europe, whereas the closely related yellow-bellied toad (B. variegata) lives in western and southern Europe. The ranges of the two species overlap in a long but very narrow zone stretching 4,800 kilometers from eastern Germany to the Black Sea (FIGURE 17.13). Hybrids between the two species suffer from a range of defects, many of which are lethal. Those hybrids that survive often have skeletal abnormalities, such as misshapen mouths, ribs that are fused to vertebrae, and a reduced number of vertebrae. By following the fates of thousands of toads from the hybrid zone, investigators found that a hybrid toad, on average, is only half as fit as a purebred individual of either species. The hybrid zone remains narrow because there is strong selection against hybrids and because adult toads do not move over long distances. The zone has persisted for hundreds of years, however, because individuals of both species continue to move short distances into it, continually replenishing the hybrid population.
355
Reproductive isolation is reinforced when diverging species come into contact
As shown in Figure 17.9, the leopard frogs Rana berlandieri and R. sphenocephala usually have nonoverlapping breeding seasons in areas of sympatry, but where the species are allopatric, both species breed in both spring and fall. But when new ponds are created where the ranges of the two species come close together, frogs from previously allopatric populations may colonize the new ponds and hybridize during their overlapping breeding seasons.
Imagine you have collected and tabulated data on hybridization between these two frog species. You have sampled various life stages of frogs and their tadpoles for 2 years after an initial spring breeding season at a newly established pond. Use the data (above right) to answer the following questions.
- Create four pie charts (one for each life stage) showing the percentage of each species and the percentage of hybrids at each stage.
- What are some possible reasons for the differences in the percentages of hybrids found at each life stage? Suggest some postzygotic isolating mechanisms that are consistent with your data.
- Over time, what changes might you expect in the breeding seasons of the two species at this particular pond, and why? How would future pie charts change if your predictions about breeding seasons are correct?

CHECKpointCONCEPT17.4
- Why would reinforcement of prezygotic isolating mechanisms be expected even if postzygotic isolating mechanisms already exist?
- Why would you expect to find different types of prezygotic isolating mechanisms among different groups of organisms? Describe some specific examples.
- Why don’t most narrow hybrid zones, such as the one between Bombina bombina and B. variegata, get wider over time?
The result of 4 billion of years of evolution has been many millions of species, each adapted to live in a particular environment and to use environmental resources in a particular way. In the next chapter we will describe how geological and biological processes interacted over the course of Earth’s history to produce this vast biodiversity.
Can biologists study the process of speciation in the laboratory?
356
ANSWER Although speciation usually takes thousands or millions of years, and although it is typically studied in natural settings such as Lake Malawi, some aspects of speciation can be studied and observed in controlled laboratory experiments. Most such experiments use organisms with short generation times, in which evolution is expected to be relatively rapid.
William Rice and George Salt conducted an experiment in which fruit flies were allowed to choose food sources in different habitats, where mating also took place. The habitats were vials in different parts of an experimental cage (FIGURE 17.14). The vials differed in three environmental factors: (1) light; (2) the direction (up or down) in which the fruit flies had to move to reach food; and (3) the concentrations of two aromatic chemicals, ethanol and acetaldehyde. In just 35 generations, the two groups of flies that chose the most divergent habitats had become reproductively isolated from each other, having evolved distinct preferences for the different habitats.

The experiment by Rice and Salt demonstrated an example of habitat isolation evolving to function as a prezygotic isolating mechanism. Even though the different habitats were in the same cage, and individual fruit flies were capable of flying from one habitat to the other, habitat preferences were inherited by offspring from their parents, and populations from the two divergent habitats did not interbreed. Similar habitat selection is thought to have resulted in the early split in cichlids that preferred the rocky versus the sandy shores of Lake Malawi.
In controlled experiments like this one, biologists can observe many aspects of the process of speciation directly.