CONCEPT37.1 Sexual Reproduction Depends on Gamete Formation and Fertilization
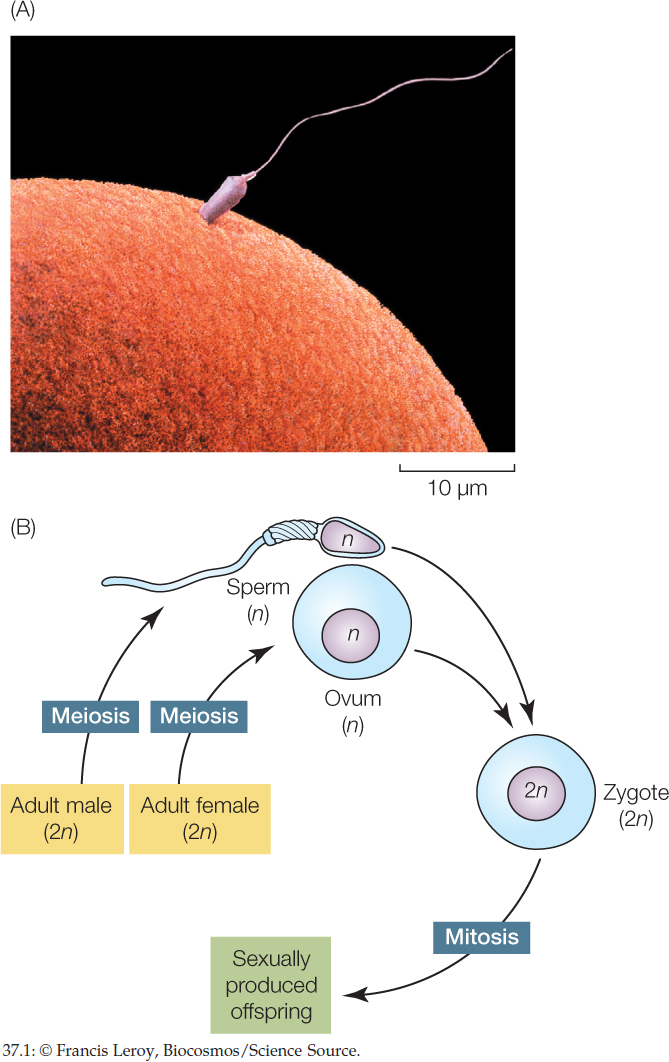
In casual conversation, “having sex” refers to human sexual activity. Biologists, however, mean something far broader when speaking of sex. As applied to animals, the process of sex is a mechanism by which the genes of two individuals are combined to produce offspring. In the case of animal sexual reproduction, each parent produces specialized reproductive cells called gametes by meiosis. Because the gametes are products of meiosis, each gamete cell has only half as many chromosomes as the other cells in the body, and the gametes are diversified in their chromosomes and genes by independent assortment and crossing over (see Concept 7.4). The gametes of the female parent are relatively large, nonmotile cells called eggs or ova (singular, ovum). The gametes of the male, called spermatozoa or sperm, are small cells that swim, typically using flagella (FIGURE 37.1A). A sperm and egg fuse to produce a single-celled zygote that has a full set of chromosomes and develops into a sexually produced offspring (FIGURE 37.1B; see Chapter 38). In this way, chromosomes and genes from two parents are combined to produce each offspring, and the new individual is not genetically identical to either parent.
LINK
The genetic aspects of sexual reproduction and the diversity it generates are discussed in Concept 7.1 and Concept 7.4; the evolutionary consequences are discussed in Concept 15.6
During asexual reproduction, by contrast, the process of sex (combination of genetic material from two parents) does not occur. Instead, offspring are produced by just a single parent, usually by mitosis: parental cells with a full set of chromosomes divide to produce new offspring cells that each have the same full set of chromosomes. Offspring are thus genetically identical to their single parent. A variety of animals, mostly invertebrates, reproduce asexually. Usually they can also reproduce sexually. Budding and fission are the two most common types of asexual reproduction. In budding, new individuals form as outgrowths, or buds, from the bodies of other individuals. In fission, an individual splits into two or more pieces that grow into new individuals. Reef corals exemplify animals in which budding is a major mechanism of reproduction. Corals occasionally reproduce sexually with eggs and sperm. However, when a new colony is produced sexually, it starts with just one small polyp. The large reef structures with which we are familiar are produced by budding. The initial polyp produces other polyps by budding, resulting in a large colony of hundreds or thousands of genetically identical polyps (FIGURE 37.2).
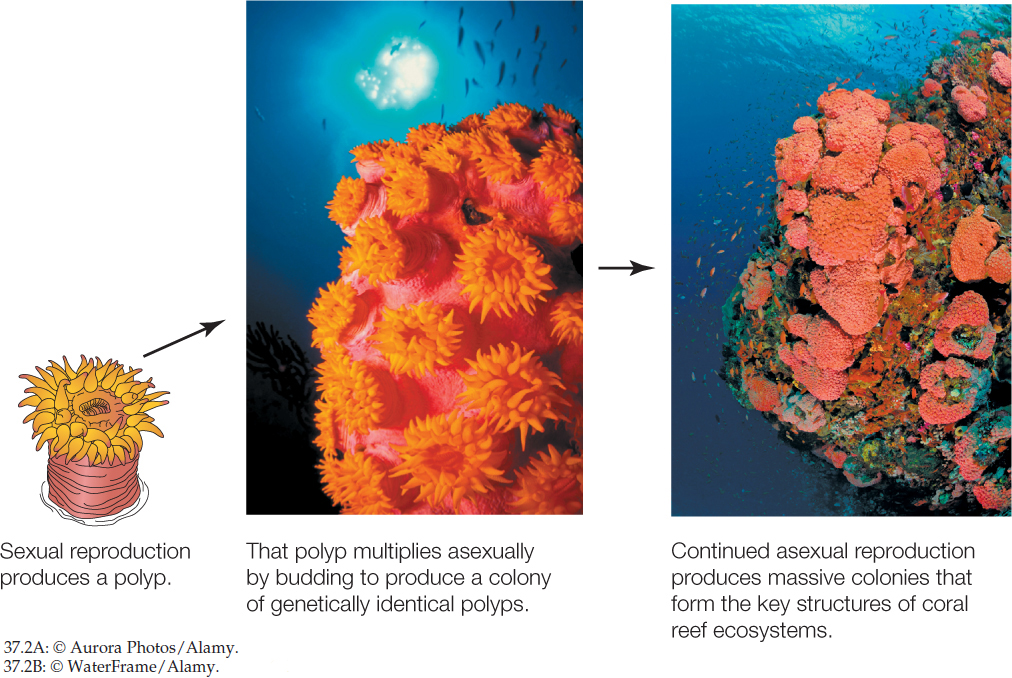
Asexual reproduction has the advantage that it maintains favorable combinations of genes. An individual with a genotype that is highly suited to its environment can rapidly produce offspring with the same advantageous genotype by budding, fission, or other asexual means.
Most animals reproduce sexually
Most vertebrates reproduce only sexually, and the same can be said of many invertebrates. Almost all animals have the capacity to reproduce sexually even if, like corals, they also reproduce asexually. We will focus on sexual reproduction for the rest of this chapter.
Sexual reproduction can seem to us to be the obvious best option for producing offspring. Why would reproduction occur by any other means? However, sexual reproduction can have some disadvantages that asexual reproduction does not (see Concept 15.6). For example, potential mates have to find each other and succeed in mating despite the presence of competitors and predators. Possibly the greatest downside of sexual reproduction is that it breaks up favorable combinations of genes. An individual with a genotype that is virtually perfect for its environment cannot simply pass that genotype along to its offspring. Biologists have theorized a great deal about why sex is so common despite this limitation (see Concept 15.6). According to one argument, the explanation lies in the changeable nature of environments. Environments, it is argued, change sufficiently—from time to time and from place to place—that no single genotype is likely to be highly successful for very long. Sexual reproduction has an enormous potential for producing genetic diversity in offspring, maximizing the odds that at least some offspring will succeed in a changing environment.
770
Among sexually reproducing animals, the two sexes, male and female, differ in three ways:
- Primary reproductive organs
- Accessory reproductive organs
- Secondary sexual characteristics
The primary reproductive organs (primary sex organs) are the gonads: the ovaries in females and testes (singular, testis)—sometimes called testicles—in males (FIGURE 37.3). These organs are “primary” because they produce the cells, the gametes, that will combine to produce a new individual.
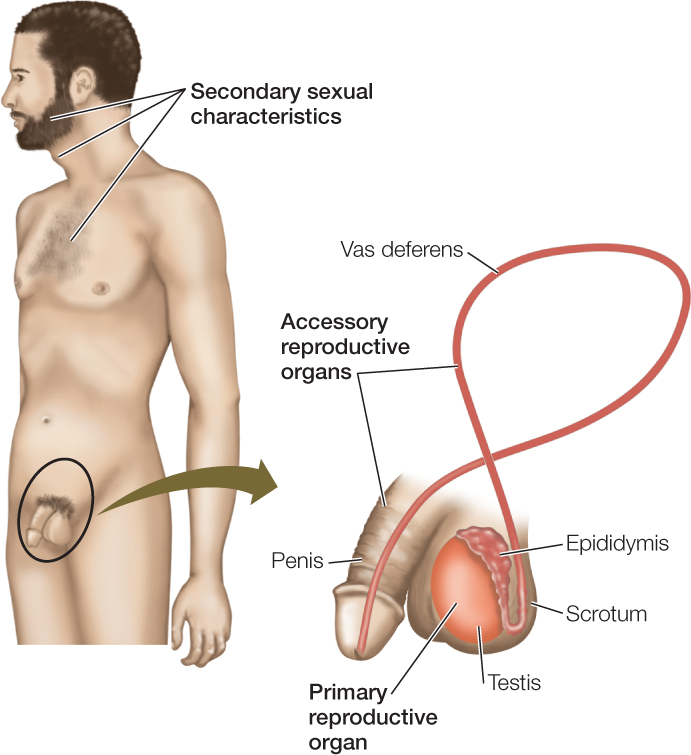
The accessory reproductive organs (accessory sex organs) are the reproductive organs other than the gonads. In human males, for example, the ducts (e.g., the vas deferens) that carry sperm out of the testes to the urethra in the penis are accessory reproductive organs, as is the penis itself. Often, the accessory reproductive organs are more elaborate in females than males. The females of some animals, such as mammals and many sharks, nurture offspring internally for a long time before giving birth. In these cases, the uterus and other structures responsible for nurturing the young are among the accessory reproductive organs. In other animals, such as birds, the females lay eggs, and their accessory organs include the glands that add the shell or capsule to each egg. (Note that the term “egg” has two meanings in discussing reproduction. The female gamete (ovum) is called an “egg,” but a structure like a hen’s egg is also called an “egg.”)
The secondary sexual characteristics are properties of nonreproductive tissues and organs that are distinctive in each sex. These characteristics often result from the actions of sex hormones on nonreproductive structures. Facial hair, chest hair, and a deep voice are some of the secondary sexual characteristics of human males (see Figure 37.3). The bright feather colors of many male birds and the enlarged horns of male caliper beetles (Golofa porteri) are also secondary sexual characteristics:
771

Secondary sexual characteristics are frequently subject to sexual selection.
LINK
Sexual selection results from nonrandom mating and is discussed in Concept 15.2; see especially Figures 15.9 and 15.10
Gametogenesis in the gonads produces the haploid gametes
Gametogenesis is the process by which gametes are produced. More specifically, it is called spermatogenesis in males and oogenesis in females. The process begins with diploid germ cells (diploid, denoted 2n, means two copies of each chromosome per cell; see Figure 37.1). These are cells in the gonads that are capable of undergoing meiosis to produce gametes. The diploid germ cells divide by mitosis until they reach a stage at which they switch to meiosis. These mitotic divisions increase the numbers of cells that are available to undergo meiosis and thereby increase the number of haploid gametes produced (haploid, 1n, means one copy of each chromosome per cell). Recall that meiosis consists of two cell divisions, meiosis I and meiosis II, during which the DNA is replicated only once (see Figure 7.11).
In adult males, sperm are typically produced continuously during the time of year when reproduction occurs (i.e., all year in human males). During this time, the populations of diploid germ cells in the testes multiply continuously, producing cells that go through the two meiotic divisions, resulting in four, equal-sized haploid cells from each diploid cell (FIGURE 37.4A). The haploid cells then mature into fully differentiated sperm.
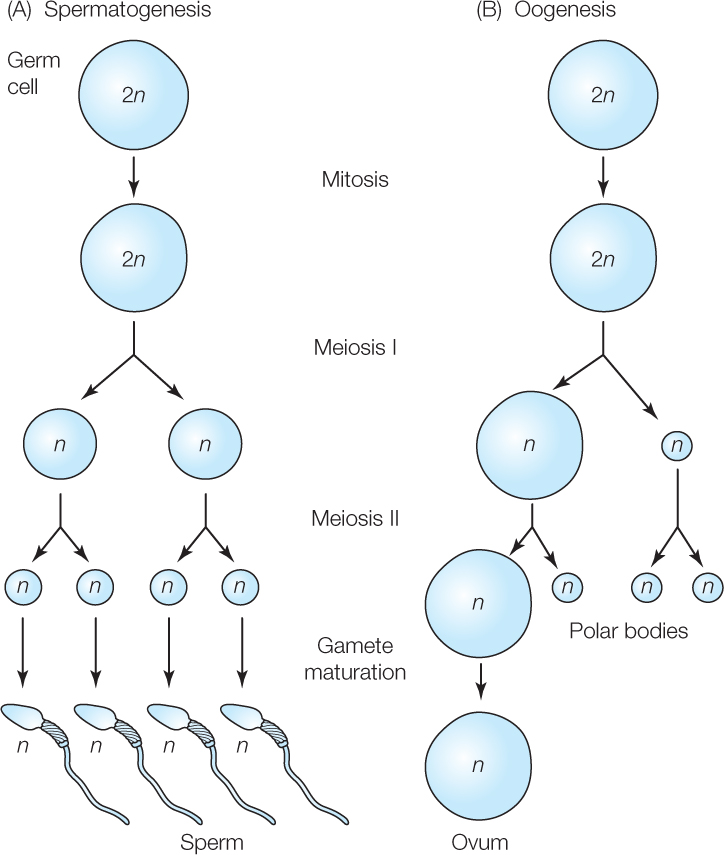
Gametogenesis in females differs in two ways from that in males. First, meiosis typically produces only one gamete from each diploid germ cell because the cytoplasm of the diploid cell is divided in highly unequal ways during each of the two meiotic divisions (FIGURE 37.4B). One of the cells produced by meiosis is a large cell, the ovum. The others are tiny cells, called polar bodies, that degenerate.
The second difference that often occurs is cellular developmental arrest during meiosis in females. When a cell destined to become an ovum is in arrest, it can continue to accumulate resources and grow, but its meiosis stops. Mammals exemplify developmental arrest during oogenesis. According to the prevailing view of most researchers, all the diploid germ cells in a young female placental mammal stop undergoing mitosis, begin meiosis (completing the very first step in meiosis I), and go into developmental arrest while the individual is a fetus in the uterus or shortly thereafter. The cells then remain in arrest for months or years until the individual matures to reproductive age. Then some of the arrested cells resume meiosis to become ova that are released from the ovaries. In humans, one of the arrested cells typically develops fully and is released from the ovaries during each month between the time a girl enters puberty (near age 10) and the time she goes through menopause (near age 50).
The gonads, in addition to having germ cells and producing gametes, typically contain somatic cells that contribute to reproduction. Somatic cells by definition do not produce gametes. If they divide, they do so strictly by mitosis. Certain somatic cells in the gonads provide metabolic support to the germ cells during gametogenesis. Somatic cells may also secrete hormones. The mammalian testis illustrates these points (FIGURE 37.5).
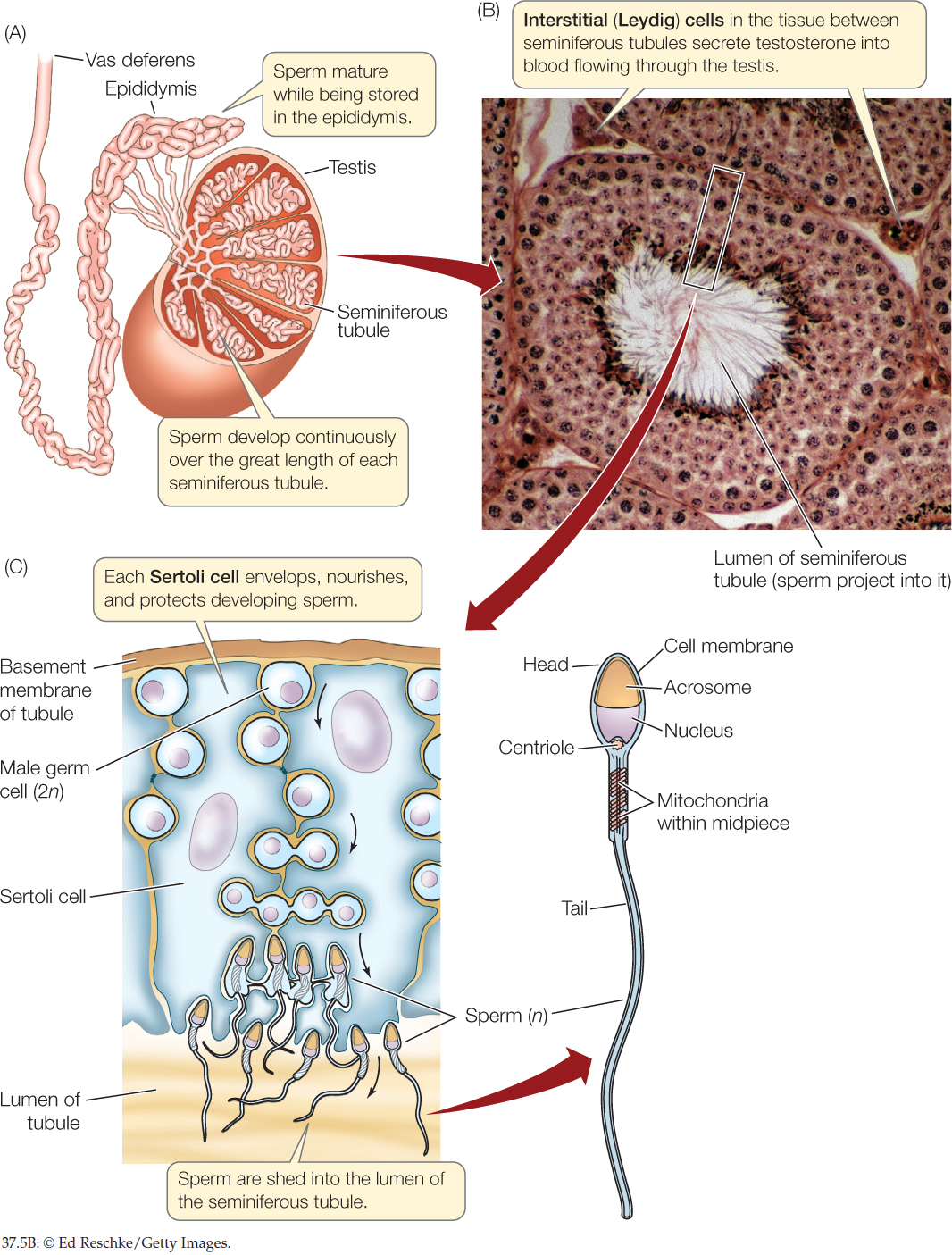
772
There are three principal cell types in a testis:
- Germ cells that produce the sperm
- Somatic Sertoli cells that assist sperm production
- Somatic interstitial cells (Leydig cells) that produce the steroid sex hormone testosterone (see Concept 35.4)
The testis is principally composed of long, coiling tubules, the seminiferous tubules. The lining of each tubule consists of germ cells in all the stages of gametogenesis and Sertoli cells. Sperm are shed into the lumen (open central cavity) of the tubule and make their way to the epididymis, a highly convoluted tubular structure attached to the outside of the testis, where the sperm undergo maturation. The epididymis connects to the vas deferens, a tube that carries sperm out of the testis during ejaculation (see Figure 37.3). The endocrine interstitial cells are located in the spaces between adjacent seminiferous tubules.
Let’s focus more closely on the somatic cells that support the germ cells during gametogenesis. In male vertebrates, spermatogenesis occurs in the midst of Sertoli cells (see Figure 37.5C). The Sertoli cells envelop the developing germ cells, provide nutrients, and help regulate sperm production. A striking fact is that the Sertoli cells constitute well over half the volume of each testis in a human or other mammal. This testifies to their importance.
Ova often have a more defined relationship with somatic support cells, in the sense that each ovum is associated with a specific set of support cells during its development. This is true in vertebrates and some invertebrates (e.g., insects). A developing ovum and its support cells are together called an ovarian follicle. In human females, as we’ve said, one cell that entered developmental arrest during fetal life typically develops to become an ovum released from the ovaries during each month of adult life. The developing ovum, termed an oocyte, is surrounded by somatic support cells—termed granulosa and theca cells—that multiply as development takes place (FIGURE 37.6). As the support cells multiply, they provide increased metabolic support, and the follicle grows enormously from a primodial follicle to a mature (Graafian) follicle. Along the way, a fluid-filled cavity (antrum) opens up within it. Moreover, the follicle migrates to be at the very surface of the ovary, where at full maturity it bulges out like a large blister, 1.5–2 centimeters in diameter in a human. The release of an ovum from the ovary is called ovulation. In vertebrates, including humans, it occurs by rupture of the mature follicle.
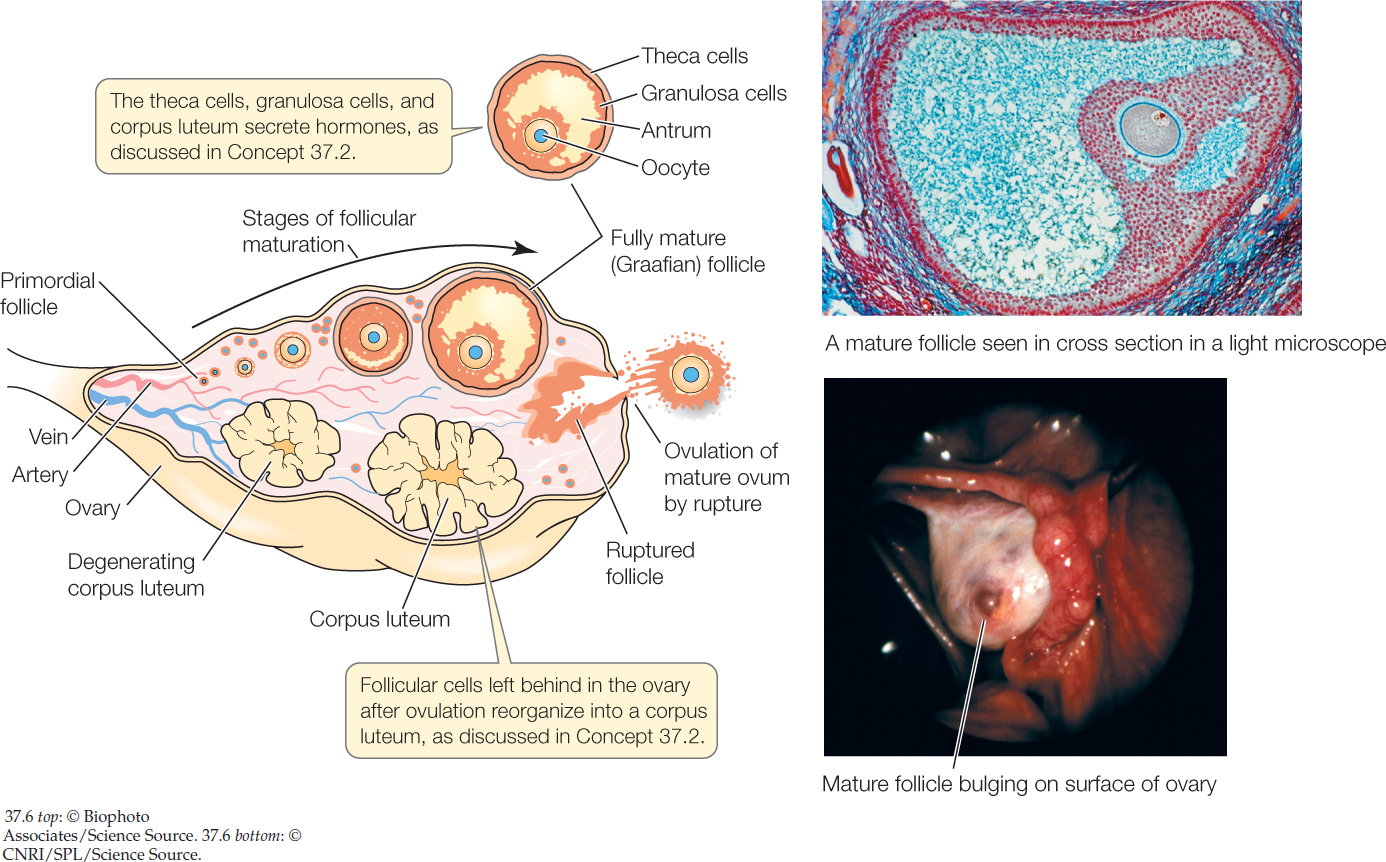
773
Fertilization may be external or internal
Fertilization is the fusion of sperm and ovum. It can be external or internal. The product is a single diploid cell, the zygote, which develops into an embryo (see Chapter 38).
To achieve fertilization, most aquatic animals simply release their gametes into the water, a process termed spawning. Fertilization then occurs in the water outside the female’s body and is described as external fertilization. In spawning species, the ducts and openings of the reproductive system can be similar in males and females because both sexes are simply releasing gametes into the environment. The zygotes typically do not receive parental care. They must develop into feeding larvae and find food before the yolk or other nutrients provided in the ovum run out. Mass deaths of commercially important fish larvae sometimes occur when the larvae are only 2–3 weeks old because at that time the larvae exhaust the yolk provided in the eggs but fail to find food. With the larvae dead, adults are not produced.
Internal fertilization occurs when a male inserts sperm inside the reproductive tract of a female, permitting fertilization to occur inside the female’s body. Most terrestrial animals employ internal fertilization—not surprising, because it ensures that the gametes and zygotes are kept moist. Some aquatic animals also employ internal fertilization.
Internal fertilization can have several advantages. It permits parental defense of the new zygotes. Some animals enclose each fertilized egg in a protective shell or capsule before laying the egg. This is true in birds (e.g., chicken eggs), insects, turtles, skates, and some sharks. Internal fertilization is essential for this process because sperm must reach the ovum before the shell is formed:

Some animals—notably mammals and certain sharks (e.g., requiem and hammerhead sharks)—keep fertilized zygotes in the female reproductive tract for an extended period of development during which nutrients are steadily provided, so the young become large and developmentally advanced before birth. Another advantage of having the male insert sperm in the female reproductive tract is that the female can potentially store sperm for future use in fertilization (see Concept 37.3).
774
What are some of the ways that sperm are introduced into the female reproductive tract for internal fertilization? Mammals have a penis that is stiffened by an internal bone or by blood pressure (see Concept 37.2). Insects and some snails also have penises. Sharks have a specialized, penis-shaped fin element (clasper) with a groove. The clasper is inserted in the female genital opening, and sperm are washed down the groove into the female reproductive tract. Crabs assemble sperm into packets called spermatophores that they grasp with modified legs and insert. Squids and octopuses insert spermatophores by use of specialized tentacles. Birds achieve sperm transfer without a penis or similar organ, merely by the act of the male and female touching their cloacal openings together (a “cloacal kiss”).
To achieve fertilization, a key challenge that must be met is for sperm and eggs of the same species to be brought together. This process involves both species recognition and synchronization.
Animal behavior often plays a major role in bringing sperm and eggs of the same species together. Males and females associate in species-specific ways (see Concept 40.6). They often also employ behaviors to signal reproductive readiness. For example, the females of many spontaneously ovulating species of mammals enter a sexually receptive behavioral condition called estrus, or “heat,” only at times when there is a high likelihood that they are releasing ova from their ovaries (see Concept 37.2). Otherwise they ignore or repel males.
Important mechanisms also exist at the cellular level for bringing sperm of a species together with eggs of the same species. Ova often attract sperm with species-specific chemical attractants.
When sperm and ova meet, cellular mechanisms perform several functions. There are cellular mechanisms that prevent fertilization by sperm of the wrong species. Such recognition mechanisms are particularly important in aquatic species that practice external fertilization because the sperm and ova of different species mix in the aquatic environment. There are also cellular mechanisms that ensure that only a single sperm enters an ovum, regardless of how many sperm may be trying. These mechanisms are termed blocks to polyspermy.
Sea urchin ova, which have been studied to an exceptional extent, illustrate these mechanisms. To fertilize an ovum, an urchin sperm must first get through two protective layers that surround the ovum—a jelly coat and a proteinaceous vitelline envelope. FIGURE 37.7 summarizes the successive mechanisms that ensure the sperm and ovum are of the same species, permit the sperm to reach the ovum, and block polyspermy. The cell membrane of the sperm ultimately fuses with that of the ovum, permitting the nucleus of the sperm to enter the ovum so a diploid zygote is formed.
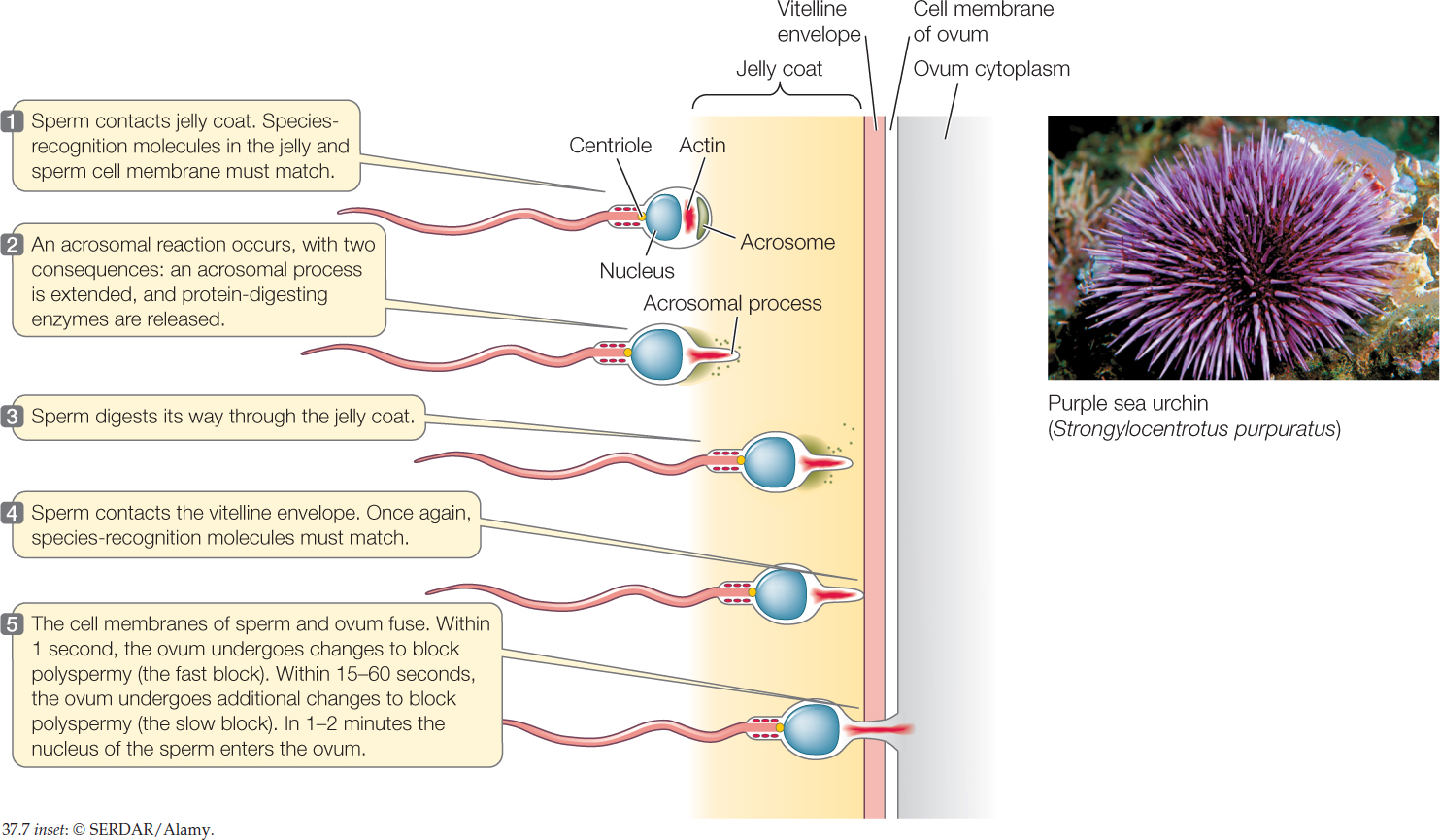
The sex of an offspring is sometimes determined at fertilization
Sex determination is the process by which the sex of an individual becomes fixed. Sex determination is not understood in all animals, but among the animals in which it is understood, a wide variety of mechanisms are known.
775
Sex is determined at fertilization in humans and other placental mammals. This was first analyzed through studies of chromosomes. One pair of the chromosomes in a mammal is distinctive because the two chromosomes in the pair differ greatly in size in males. These are called the sex chromosomes. The larger and smaller are named X and Y, respectively. Human cells have 23 pairs of chromosomes. The two chromosomes in 22 of these pairs are always similar to each other in their appearance under a light microscope, regardless of whether an individual is male or female (see Chapter 7, Concept 7.2). The 23rd pair consists of an X and a Y chromosome (in males) or two X chromosomes (in females).
Because adult females have two X chromosomes, during meiosis all ova receive an X chromosome. By contrast, the pair of sex chromosomes in a male consists of an X and a Y. Thus as sperm are produced, half receive an X chromosome and half receive a Y.
A human zygote’s sex is determined at fertilization by the sex chromosomes it receives. It receives an X from its mother but may receive either an X or a Y from its father. A zygote that receives an X from both parents is XX and female. One that receives an X and a Y is XY and male.
Long after this chromosomal explanation of sex determination was known, researchers discovered a particularly critical gene that they named SRY (sex-determining region of the Y chromosome). This one gene exerts exceptional influence on sexual development, although many other genes are involved. The gene is nearly always found on the Y chromosome. An XY zygote thus has the gene, which directs the primordial gonads (the gonads early in development) to become testes and secrete testosterone. An XX zygote lacks the SRY gene and develops into a female. Concept 35.4 and Figure 35.13 discuss how testosterone secretion in XY embryos leads to phenotypic sex differentiation.
Fruit flies (Drosophila) also have X and Y chromosomes (see Concept 8.3). Their mechanism of sex determination is very different, however, because the Y chromosome and its genes have no direct role.
Sex is not determined at fertilization in some animals. Of these, crocodilians and turtles are of particular interest because many species exhibit environmental sex determination. The sex of each individual is determined by the temperature it experiences during its embryonic development. American alligators, for example, become males if they develop at 32–33°C, have a 50:50 chance of becoming male or female at 34°C, and develop into females at 35°C. Great worry exists that these animals will be exceptionally vulnerable to global warming because temperatures that are higher than traditional norms may lead to maladaptive changes in their sex ratios.
Some animals undergo sex change
Some animals do not undergo sex determination. Instead, they can be males at one time and females at another during their lives. They are known as sequential hermaphrodites. Hermaphrodites are animals that can produce both sperm and ova. In sequential hermaphrodites, sperm and ova are not produced at the same time but instead are produced at different times in an individual’s life. Sex change in these animals does not entail anything as dramatic as exchanging a penis for a vagina. The animals are aquatic and release their gametes into the water by spawning. Their accessory reproductive organs are suited to carrying either sperm or eggs to the outside. Sex change occurs in the type of gametes they produce. Great numbers of fish species, including many coral reef species, undergo sex change. The phenomenon also occurs in certain aquatic annelid worms, and in certain snails or other gastropod mollusks.
Biologists think that sex change has evolved in cases in which an individual’s reproductive output—its production of offspring—is maximized by changing sex. What conditions affect reproductive output? A great many do, including a species’ breeding system (the way in which males and females associate), social organization, and population density. Because of this complexity, many different types of sex change occur.
Anemonefish (Amphiprion percula), which live among the tentacles of a sea anemone, provide a well-studied example (FIGURE 37.8). Under many circumstances, the fish on a single sea anemone consist of one breeding male that spawns sperm, one breeding female that spawns ova, and several nonbreeding males that do not spawn. The breeding female is largest, and the breeding male is second largest. All the fish start life as males. When the female dies, the breeding male becomes the largest fish and turns into a female. One of the nonbreeding males becomes the new breeding male. According to the prevailing theory of sex change, the reason the breeding male changes to a breeding female is that he can have more offspring by being female. To see why, remember that there are only two breeding fish on an anemone. A fish of any size can produce huge numbers of sperm. If the biggest fish remained male, its reproductive output would be limited by the egg output of a smaller fish. By switching to producing eggs, the biggest fish increases the number of eggs that get fertilized and thus its reproductive output (because egg production increases with body size). Currently, investigators are starting to make progress in understanding the hormonal mechanisms of sex change (FIGURE 37.9).

Investigation
HYPOTHESIS
An increase in the concentration of 11-ketotestosterone (11-KT) is sufficient to cause female-to-male transition in blackeye gobies

- Capture female blackeye gobies in the wild and hold in individual tanks.
- Insert in each fish a hormone-releasing implant containing 11-KT or an implant with no hormone (control treatment). The implant is designed to release hormone gradually.
- After 43 days of treatment, examine the gonads of the fish to determine the sex.

CONCLUSION
The results support the hypothesis that an increase in the concentration of 11-KT is sufficient to cause female-to-male transition.
ANALYZE THE DATA
In a related experiment, 8 female fish were treated with fadrozole, a chemical that inhibits the synthesis of 17β-estradiol and thus should lower the concentration of this female sex hormone. At the end of treatment, 1 fish was still female, 5 fish were transitional, and 2 fish were male.
- Was this treatment as effective as 11-KT (or 11-ketoadrenosterone) administration in causing females to become males? What data support your conclusion?
- The researchers hypothesized that a decrease in 17-estradiol causes sex change. Do the data support the hypothesis? Why or why not? What additional information would be helpful in deciding whether the hypothesis is supported?
aF. J. Kroon and N. R. Liley. 2000. General and Comparative Endocrinology 118: 273–283.
776
CHECKpointCONCEPT37.1
- What are some of the advantages and disadvantages of internal and external fertilization?
- Vasectomy is a procedure for birth control in which the principal ducts carrying sperm out of the testes are tied off. Basing your answer on the cellular structure of the human testis, explain why a man’s testosterone level is not affected by vasectomy.
- Rarely, men are found who have two X chromosomes. The SRY gene is responsible for this syndrome. What type of unusual gene placement could account for the syndrome? Explain.
Having surveyed the general aspects of animal gametogenesis and fertilization, we will next consider the male and female reproductive systems in mammals, using our own species as the primary example.