Mutualisms can affect communities
When we think about mutualisms, we often focus on how the relationship helps each of the interacting species; it often improves the fitness and abundance of each participant. For example, corals cannot survive without zooxanthellae and many plants cannot produce offspring without pollinators. In addition to affecting abundance, mutualisms can also affect other ecological levels. In this section, we will examine how mutualisms, and the interruption of mutualisms, can alter the distributions of species, the diversity of communities, and the functioning of ecosystems.
404
Effects on Species Distributions
Given the benefits of mutualisms, we might expect natural selection to favor mutualistic relationships among species and to expand the distribution of species engaged in mutualisms. In contrast, we would expect the disruption of a mutualism to cause a decline of the species involved and a reduction in their distribution. We can see an example of this in the plant known as garlic mustard (Alliaria petiolata), a member of the mustard family with leaves that smell like garlic when crushed. Native to Europe and Asia, it was introduced to North America more than a century ago for human consumption. When garlic mustard grows in North American forests, it causes young trees to grow poorly and makes them less likely to reach adult size. For a number of years, researchers sought to understand the mechanism underlying garlic mustard’s harmful effects.
In 2006, researchers discovered that garlic mustard was interfering with the mutualism between the forest trees and arbuscular mycorrhizal fungi in the soil. To demonstrate this link, they examined how well three species of trees grew when raised in soil where garlic mustard had invaded and in soil where it had not invaded. As you can see in Figure 17.19a, the increases in biomass of sugar maple, red maple, and white ash trees were many times greater when grown in soil that had not been invaded by garlic mustard. Moreover, when the researchers examined the percentage of tree roots colonized by mycorrhizal fungi, they found that soil collected from forests with garlic mustard had little or no colonization by mycorrhizae, as illustrated in Figure 17.19b.

Additional experiments revealed that the roots of garlic mustard produce chemicals that inhibit the growth of mycorrhizal fungi. Given that tree species depend on fungal mutualism to different degrees, garlic mustard has the largest negative effect on those tree species that have the greatest dependence on the fungi. Because garlic mustard disrupts vital mutualisms, it has the potential to alter the distribution of a large number of other species as it spreads across North America.
Effects on Communities
Mutualisms that alter the abundance and distribution of one or more species can have widespread effects on the rest of the community. A community can be affected in several ways; the mutualism might cause a change in species diversity or it might alter the abundance of individuals within species in the community.
Altering Species Diversity
A generalist mutualist species interacts with many other species and offers widespread benefits. In the case of the cleaner fish, for example, a particular species of cleaner fish may remove parasites from many different species of larger fish. If this behavior helps species to persist on a coral reef, then the removal of cleaner fish should cause a decline in the total number of large fish and the number of species. Researchers working on coral reefs in Australia tested this question by removing a species of cleaner fish (Labroides dimidiatus) from nine small reefs. They designated another nine reefs as controls. After 18 months, they counted both the number of individual fish and the number of fish species on each reef. As you can see in Figure 17.20, removing the cleaner fish caused the number of individual fish to decline by about three-quarters and the number of species to decline by half, which suggests that cleaner fish play a critical role in maintaining populations of reef fish.

405
Initiating a Chain of Interactions
In some cases, the community does not lose species when a mutualism is disturbed, but the abundance of many species changes through a chain of interactions. Earlier in this chapter we discussed the role of ants in defending acacia trees against herbivores. We saw that the ants reduce herbivory on acacia trees, which improves tree survival. In return, ants benefit from having a place to make nests and a source of food in the nectaries. However, what would happen if much of the herbivory is removed and the ants no longer provide a benefit?
Researchers examined this question in the savanna region of Kenya. They set aside 12 plots of 4 ha each; half of the plots were fenced to exclude all large herbivores whereas the other half were left as unfenced controls. The plots contained 40- to 70-year old acacia trees (Acacia drepanolobum). After 10 years, researchers examined how fencing the plots affected the ant-acacia mutualism and the rest of the community. You can view their data in Figure 17.21. In the fenced plots that lacked large herbivores, trees produced fewer swollen thorns and nectaries than trees in the control plots, as shown in 17.21a and 17.21b.

406
ANALYZING ECOLOGY
Comparing Two Groups That Do Not Have Normal Distributions
As we saw in the case of the cleaner fish, researchers who study mutualisms often need to test whether the mutualistic interaction really provides a benefit to each species in the interaction. To do so, they need to conduct statistical tests that compare how each species performs both with and without the presence of the other species. In Chapter 15 we discussed the use of t-tests to compare the means of two groups. As you may recall, t-tests require that the data collected follow a normal distribution, which are the bell-shaped curves that we discussed in Chapter 2. In some cases, however, the data from two groups do not have normal distributions, so we cannot use the t-test. In these cases, we need to use the Mann-Whitney rank sum test, which is named for the statisticians who developed it.
The Mann-Whitney rank sum test begins by ranking the data—from low to high—and then taking the sum of those ranks. For example, consider the following set of data for the number of species occurring in eight coral reefs where researchers have removed the cleaner fish and eight coral reefs where researchers have allowed the cleaner fish to remain. If we were to take all 16 numbers and place them in order from the lowest value to the highest value, we could give each value a rank from 1 to 16. Whenever we have more than one occurrence of a value, we assign both values the average rank.
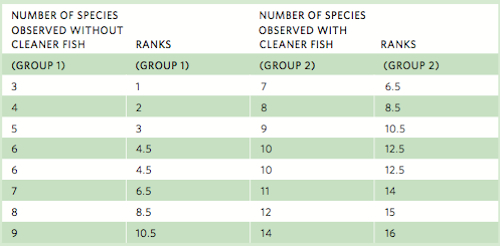
The next step is to sum the ranks for group 1 (denoted as R1) and group 2 (denoted as R2).
R1 = 1 + 2 + 3 + 4.5 + 4.5 + 6.5 + 8.5 + 10.5 = 40.5
R2 = 6.5 + 8.5 + 10.5 + 12.5 + 12.5 + 14 + 15 + 16 = 95.5
We can then use either value of R to calculate a test statistic known as U. We will use R2:
U = R2 − (n2 × (n2 + 1) ÷ 2)
where n1 is the number of observations in group 1 and n2 is the number of observations in group 2. Using this formula,
U = 95.5 − (8 × (8 + 1) ÷ 2) = 59.5
Now that we know the value of U, we need to calculate the mean and standard deviation of U for the entire collection of data from both groups. The mean value of U is:
mU = (n1 n2) ÷ 2
mU = (8 × 8) ÷ 2 = 32
The standard deviation of U is:
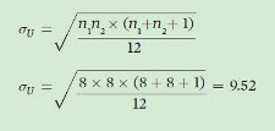
Using our calculated values of U, mU, and σU we can calculate z, which is defined as:

We can look up this value in a table of z values in the Statistical Tables appendix, where we find that the probability value is 0.002. Since this probability is less than 0.05, we can conclude that the groups are significantly different from each other. This means that removing the cleaner fish causes a significant decrease in the number of fish species that live on a coral reef.

U = R1 − (n1 × (n1 + 1) ÷ 2)

Changes in the acacia trees caused subsequent changes in the abundances of mutualistic ants. One species of ant, Crematogaster mimosa, relies heavily on the swollen thorns for making its nests and raising its offspring. The reduction of swollen thorns in the fenced plots caused a 30 percent reduction in the proportion of trees occupied by this ant, as illustrated in Figure 17.21c. Moreover, of those trees that were occupied, the average colony of C. mimosa was 47 percent smaller than in the control plots.
Another ant species, C. sjostedti, does not use the swollen thorns for nests but instead nests in tree cavities that are excavated in the acacia tree by long-horned beetles. Because C. sjostedti does not require swollen thorns, the decline in C. mimosa in the fenced plots allowed C. sjostedti to double the proportion of trees that it occupied. This increase in C. sjostedti had further effects on the community. In contrast to C. mimosa, which works to eliminate insect herbivores, C. sjostedti allows long-horned beetles to live on the trees and bore holes into the trunk as it slowly consumes the trees. The beetle receives a food benefit and C. sjostedti receives a nest benefit, so these two species represent another mutualism in the community. However, beetle excavation causes the acacia tree to grow more slowly and die at about twice the rate as trees that are tended by C. mimosa. As a result, although larger mammal herbivores can graze the acacia trees and have negative effects on tree growth, the exclusion of large herbivores turns out to have a much larger negative effect on the trees because the trees are no longer defended from competitors or long-horned beetles. As this example demonstrates, mutualisms affect more than the species involved in the interactions; they can have far-reaching effects throughout the entire community.
407
Effects on Ecosystem Function
Mutualisms can also have ecological effects at the level of the ecosystem. As we discussed in Chapter 1, researchers working on ecosystems examine the movement of energy and matter among many sources including both the biotic and abiotic realms. For example, researchers working with arbuscular mycorrhizal fungi investigated how a grassland ecosystem of 15 species would respond to different numbers of fungal species in the soil. When they examined the total amount of phosphorus that the plants took up from the soil, they found that plants living in soils that contained more fungal species took up a greater amount of phosphorus, as shown in Figure 17.22a. They also quantified the total biomass of roots in the soil and shoots above the soil. They found that a higher number of fungal species in the soil caused substantial increases in the biomass of the roots and shoots in the ecosystem. You can see these data in Figure 17.22b and Figure 17.22c. As illustrated in this example, mutualisms not only affect individual species, but they can also have large effects on the functioning of ecosystems.

408
Throughout this chapter, we have seen how species can interact in a variety of mutualisms. These interactions fulfill a wide range of needs including resource acquisition, places to live, defense, pollination, and dispersal. These interactions can have important effects on communities and ecosystems and they can even have major conservation implications, as you will see in the “Ecology Today” feature that follows.
409
ECOLOGY TODAY CONNECTING THE CONCEPTS
DEALING WITH THE DEATH OF DISPERSERS

The Republic of Mauritius provides an example of how important mutualisms can be to the persistence of species in nature. Mauritius, a group of islands located in the Indian Ocean southeast of the African continent, is the only place on Earth where the dodo bird (Raphus cucullatus) once lived. A large, flightless bird, the dodo was an easy source of food for the sailors who visited the islands, and was also harmed by the introduction of many non-native species. By the late 1600s, about 200 years after humans first set foot on Mauritius, the dodo was extinct. Numerous other species unique to Mauritius have also been driven extinct, including two species of giant tortoises: the high-backed tortoise (Cylindraspis triserrata) and the domed tortoise (C. inepta). Not only are the islands missing these species, they are also missing the services that these species provided.
Over subsequent centuries, as humans settled the islands, they cut down large swaths of tropical forest for lumber, firewood, and space for sugarcane fields. Today, only about 2 percent of the original forest remains and some of the tree species continue to decline. For example, Syzygium mamillatum is a critically endangered tree species that consists of just over a hundred individuals. Scientists studying this problem realized that many declining tree species had probably once relied on the dodos and tortoises for seed dispersal. These species consumed the fruit, and the seeds were released after they passed through the animals’ digestive systems. When the animals defecated, typically far away from the trees that produced the fruits, the seeds were ready to germinate. With these mutualists extinct, fewer fruits were consumed and any released seeds remained close to the parent tree. Seeds that geminate near the parent tree compete with the parent and there are high rates of disease among conspecific trees concentrated in one place. Without dispersers, the seeds of the rare trees cannot disperse and recolonize open areas that have been logged. Researchers wondered if it was possible to bring back the process of seed dispersal even without the animals that originally did the dispersing.
Researchers came up with a radical idea. They hypothesized that perhaps non-native species could be introduced to fill the role of the lost mutualists and help save the endangered trees. To play the role of the dodo, they brought in domestic turkeys (Meleagris gallopavo), which—similar to dodos—have a large gizzard that grinds the fruits they swallow. The researchers hypothesized that when the dodo consumed the fruits, the grinding of the gizzard broke down the fruits and released the seeds, but maintained the viability of the seeds as they passed through the bird’s digestive system. To play the role of the two species of extinct tortoises, they brought in giant Aldabra tortoises (Aldabrachelys gigantea) from the nearby Seychelles Islands.
410
To test the potential for these species to act as substitute seed dispersers, the researchers fed fruits of the S. mamillatum tree to the turkeys and Aldabra tortoises. When fruits were fed to the turkeys, the large gizzards not only broke apart the fruit but also ground the seeds into small pieces, which meant that turkeys were not suitable replacements for the dodos. However, when fruits of the tree were fed to the Aldabra tortoises, the researchers found that 16 percent of the seeds inside the fruits remained whole in tortoise feces. Although these seeds had lower germination success than seeds that did not pass through the tortoise’s gut, they produced seedlings that grew taller and had more leaves. This meant that the Aldabra tortoises might serve as an effective substitute disperser.
In a follow-up study, the researchers tested whether the tortoises could serve as a new mutualist for critically endangered trees if they were released into the wild. To test this idea, they decided to introduce Aldabra tortoises to a small, 25-ha island that is part of the Republic of Mauritius. This island contains another rare tree species, the ebony tree (Diospyros egrettarum), which was once abundant and produces large, 16-g fruits. Logging for firewood had made this tree rare on the island and large areas contained no new ebony tree seedlings. Without dispersers, the fruits landed near the parent tree and stayed there. After introducing the tortoises into pens in 2000, the researchers confirmed that the tortoises had no negative effects on the plant community. In 2005, 11 tortoises were allowed to roam freely throughout the island. In 2011, the investigators reported some remarkable results. Few fruits were now found near the parent trees because the tortoises were consuming most of them and, because the seeds passed through the tortoises when they defecated, the seeds were relocated to many places around the island. New seedlings showed up everywhere, including open areas where the tree had not existed for many decades. Moreover, ebony seeds that passed through the tortoise gut germinated better than unconsumed seeds.
After more than a decade of research, these results suggest that, although we cannot bring back extinct species, we may be able to substitute some species to resurrect mutualisms so that the original extinctions do not cause subsequent extinctions of their mutualistic partners.

SOURCES: Hansen, D. M., et al. 2008. Seed dispersal and establishment of endangered plants on oceanic islands: The Janzen-Connell model, and the use of ecological analogues. PLOS One 3: 1–13.
Griffiths, C. J., et al. 2011. Resurrecting extinct interactions with extant substitutes. Current Biology 21: 1–4.