23.4Ammonium Ion Is Converted into Urea in Most Terrestrial Vertebrates
Ammonium Ion Is Converted into Urea in Most Terrestrial Vertebrates
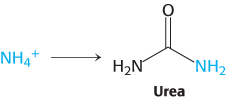
Some of the NH4+ formed in the breakdown of amino acids is consumed in the biosynthesis of nitrogen compounds. In most terrestrial vertebrates, the excess NH4+ is converted into urea and then excreted. Such organisms are referred to as ureotelic.
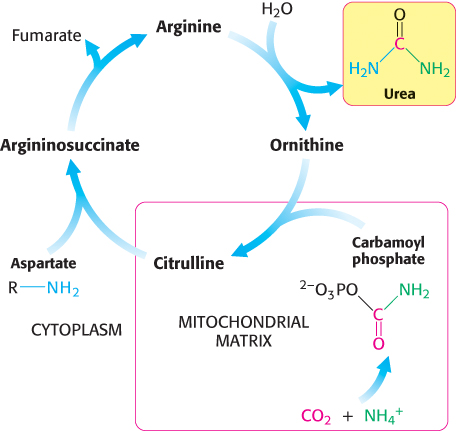
In terrestrial vertebrates, urea is synthesized by the urea cycle (Figure 23.15). One of the nitrogen atoms of urea is transferred from an amino acid, aspartate. The other nitrogen atom is derived directly from free NH4+, and the carbon atom comes from HCO3− (derived by the hydration of CO2; Section 9.2).
The urea cycle begins with the formation of carbamoyl phosphate
The urea cycle begins with the coupling of free NH3 with HCO3− to form carbamoyl phosphate, the committed reaction of the urea cycle, which is catalyzed by carbamoyl phosphate synthetase I. Carbamoyl phosphate is a simple molecule, but its synthesis is complex, requiring three steps.
694
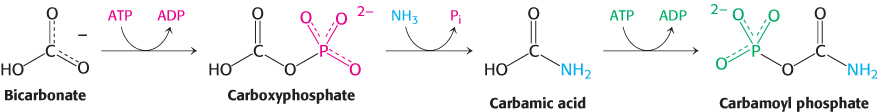
Note that NH3, because it is a strong base, normally exists as NH4+ in aqueous solution. However, carbamoyl phosphate synthetase uses only NH3 as a substrate. The reaction begins with the phosphorylation of HCO3− to form carboxyphosphate, which then reacts with NH3 to form carbamic acid. Finally, a second molecule of ATP phosphorylates carbamic acid to form carbamoyl phosphate. The structure and mechanism of the enzyme that catalyzes these reactions will be presented in Chapter 25. The consumption of two molecules of ATP makes this synthesis of carbamoyl phosphate essentially irreversible.
Carbamoyl phosphate synthetase is the key regulatory enzyme for urea synthesis
Carbamoyl phosphate synthetase is regulated both allosterically and by covalent modification so that it is maximally active when amino acids are being metabolized for fuel use. The allosteric regulator N-
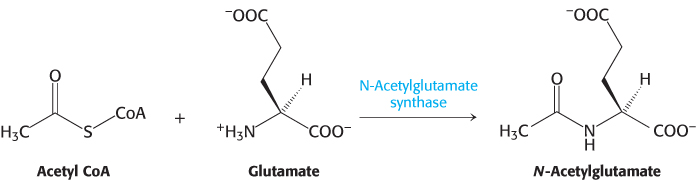
N-acetylglutamate synthase is itself activated by arginine. Thus, NAG is synthesized when amino acids, as represented by arginine and glutamate, are readily available, and carbamoyl phosphate synthetase is then activated to process the generated ammonia. When ammonia is not being generated, the synthetase is inhibited by acetylation. A rise in the mitochondrial NAD+, indicative of an energy-
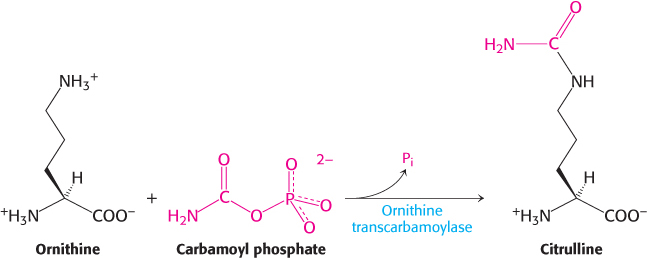
Carbamoyl phosphate reacts with ornithine to begin the urea cycle
The carbamoyl group of carbamoyl phosphate has a high transfer potential because of its anhydride linkage. The carbamoyl group is transferred to ornithine to form citrulline, in a reaction catalyzed by ornithine transcarbamoylase.
695
Ornithine and citrulline are amino acids, but they are not used as building blocks of proteins. The formation of NH4+ by glutamate dehydrogenase, its incorporation into carbamoyl phosphate as NH3, and the subsequent synthesis of citrulline take place in the mitochondrial matrix. In contrast, the next three reactions of the urea cycle, which lead to the formation of urea, take place in the cytoplasm.
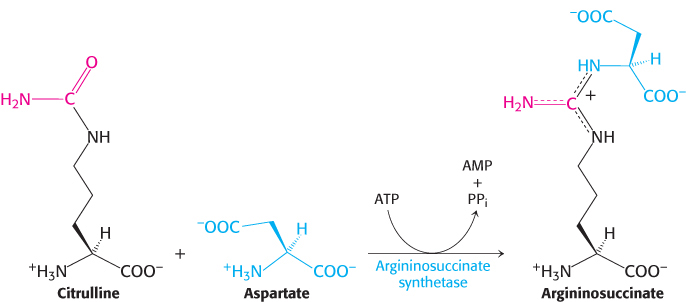
Citrulline is transported to the cytoplasm, where it condenses with aspartate, the donor of the second amino group of urea. This synthesis of argininosuccinate, catalyzed by argininosuccinate synthetase, is driven by the cleavage of ATP into AMP and pyrophosphate and by the subsequent hydrolysis of pyrophosphate.
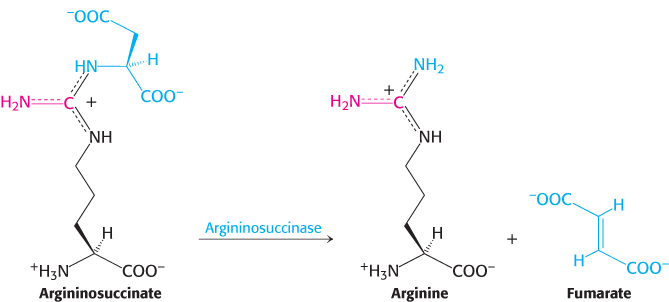
Argininosuccinase (also called argininosuccinate lyase) cleaves arginino-
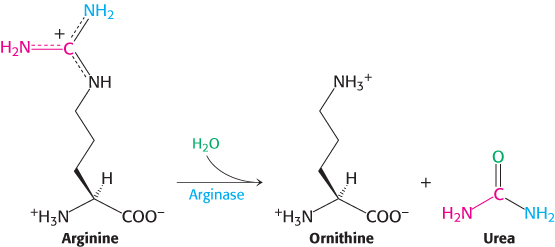
Finally, arginine is hydrolyzed to generate urea and ornithine in a reaction catalyzed by arginase. Ornithine is then transported back into the mitochondrion to begin another cycle. The urea is excreted. Indeed, human beings excrete about 10 kg (22 pounds) of urea per year.
In ancient Rome, urine was a valuable commodity. Vessels were placed on street corners for passersby to urinate into. Bacteria would degrade the urea, releasing ammonium ion, which would act as a bleach to brighten togas.
696
The urea cycle is linked to gluconeogenesis
The stoichiometry of urea synthesis is
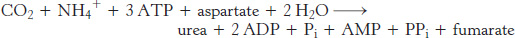
Pyrophosphate is rapidly hydrolyzed, and so the equivalent of four molecules of ATP are consumed in these reactions to synthesize one molecule of urea. The synthesis of fumarate by the urea cycle is important because it is a precursor for glucose synthesis (Figure 23.16). Fumarate is hydrated to malate, which is in turn oxidized to oxaloacetate. Oxaloacetate can be converted into glucose by gluconeogenesis or transaminated to aspartate.
Urea-cycle enzymes are evolutionarily related to enzymes in other metabolic pathways
 Carbamoyl phosphate synthetase generates carbamoyl phosphate for both the urea cycle and the first step in pyrimidine biosynthesis (Section 25.1). In mammals, two distinct isozymes of the enzyme are present. Carbamoyl phosphate synthetase II, used in pyrimidine biosynthesis, differs in two important ways from its urea-
Carbamoyl phosphate synthetase generates carbamoyl phosphate for both the urea cycle and the first step in pyrimidine biosynthesis (Section 25.1). In mammals, two distinct isozymes of the enzyme are present. Carbamoyl phosphate synthetase II, used in pyrimidine biosynthesis, differs in two important ways from its urea-
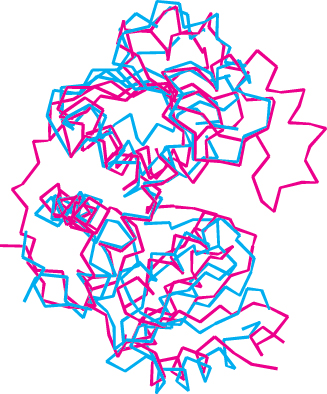
Can we find homologs for the other enzymes in the urea cycle? Ornithine transcarbamoylase is homologous to aspartate transcarbamoylase, which catalyzes the first step in pyrimidine biosynthesis, and the structures of their catalytic subunits are quite similar (Figure 23.17). Thus, two consecutive steps in the pyrimidine biosynthetic pathway were adapted for urea synthesis. The next step in the urea cycle is the addition of aspartate to citrulline to form argininosuccinate, and the subsequent step is the removal of fumarate. These two steps together accomplish the net addition of an amino group to citrulline to form arginine. Remarkably, these steps are analogous to two consecutive steps in the purine biosynthetic pathway (Section 25.2).
697
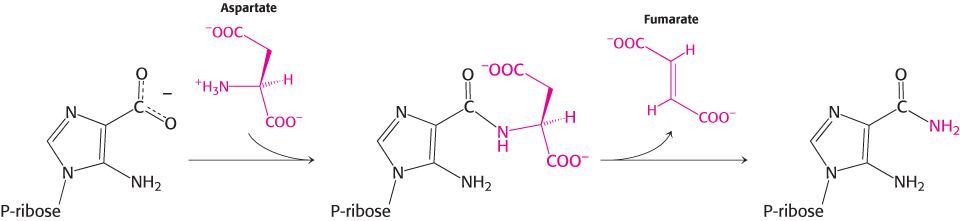
The enzymes that catalyze these steps are homologous to argininosuccinate synthetase and argininosuccinase, respectively. Thus, four of the five enzymes in the urea cycle were adapted from enzymes taking part in nucleotide biosynthesis. The remaining enzyme, arginase, appears to be an ancient enzyme found in all domains of life.
Inherited defects of the urea cycle cause hyperammonemia and can lead to brain damage
 The synthesis of urea in the liver is the major route for the removal of NH4+. Urea cycle disorders occur with a prevalence of about 1 in 15,000. A blockage of carbamoyl phosphate synthesis or of any of the four steps of the urea cycle has devastating consequences because there is no alternative pathway for the synthesis of urea. All defects in the urea cycle lead to an elevated level of NH4+ in the blood (hyperammonemia). Some of these genetic defects become evident a day or two after birth, when the afflicted infant becomes lethargic and vomits periodically. Coma and irreversible brain damage may soon follow, a condition called hepatic encephalopathy. Why are high levels of NH4+ toxic? The answer to this question is not yet known. Recent work, however, suggests that NH4+ may inappropriately activate a sodium-
The synthesis of urea in the liver is the major route for the removal of NH4+. Urea cycle disorders occur with a prevalence of about 1 in 15,000. A blockage of carbamoyl phosphate synthesis or of any of the four steps of the urea cycle has devastating consequences because there is no alternative pathway for the synthesis of urea. All defects in the urea cycle lead to an elevated level of NH4+ in the blood (hyperammonemia). Some of these genetic defects become evident a day or two after birth, when the afflicted infant becomes lethargic and vomits periodically. Coma and irreversible brain damage may soon follow, a condition called hepatic encephalopathy. Why are high levels of NH4+ toxic? The answer to this question is not yet known. Recent work, however, suggests that NH4+ may inappropriately activate a sodium-
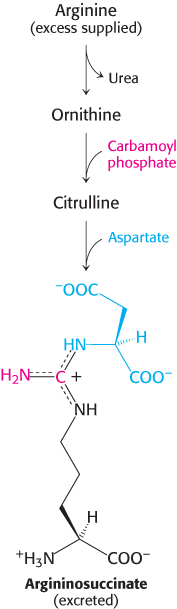
Ingenious strategies for coping with deficiencies in urea synthesis have been devised on the basis of a thorough understanding of the underlying biochemistry. Consider, for example, argininosuccinase deficiency. This defect can be partly bypassed by providing a surplus of arginine in the diet and restricting the total protein intake. In the liver, arginine is split into urea and ornithine, which then reacts with carbamoyl phosphate to form citrulline (Figure 23.18). This urea-
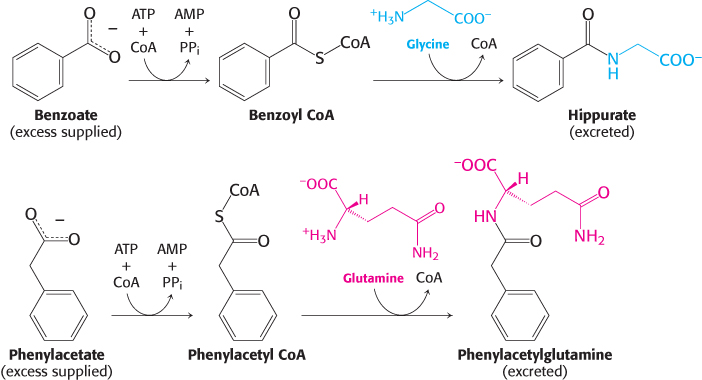
The treatment of carbamoyl phosphate synthetase deficiency or ornithine transcarbamoylase deficiency illustrates a different strategy for circumventing a metabolic block. Citrulline and argininosuccinate cannot be used to dispose of nitrogen atoms because their formation is impaired. Under these conditions, excess nitrogen accumulates in glycine and glutamine. The challenge then is to rid the body of the nitrogen accumulating in these two amino acids. That goal is accomplished by supplementing a protein-
698
Urea is not the only means of disposing of excess nitrogen
As stated earlier, most terrestrial vertebrates are ureotelic; they excrete excess nitrogen as urea. However, urea is not the only excretable form of nitrogen. Ammoniotelic organisms, such as aquatic vertebrates and invertebrates, release nitrogen as NH4+ and rely on the aqueous environment to dilute this toxic substance. Interestingly, lungfish, which are normally ammoniotelic, become ureotelic in time of drought, when they live out of the water.
Both ureotelic and ammoniotelic organisms depend on sufficient water, to varying degrees, for nitrogen excretion. In contrast, uricotelic organisms, such as birds and reptiles, secrete nitrogen as the purine uric acid. Uric acid is secreted as an almost solid slurry requiring little water. The secretion of uric acid also has the advantage of removing four atoms of nitrogen per molecule. The pathway for nitrogen excretion developed in the course of evolution clearly depends on the habitat of the organism.