Bacterium, Escherichia coli
Escherichia coli is a single-
E. coli is not a eukaryote and thus has limited homology to humans. This restricts its usefulness in studying all but the most basic cellular processes. For example, E. coli lacks nucleosomes, and most of its proteins are unmodified. Eukaryotic protein expression in transformed E. coli sometimes does not work, because it requires posttranslational modifications that the bacterium cannot carry out.
Early Studies of E. coli as a Model Organism
The marriage of biochemistry and genetics made E. coli a rich source for the discovery of new information. The famous “fluctuation analysis” by Salvadore Luria and Max Delbruck in 1943 demonstrated that E. coli mutates spontaneously to become resistant to bacteriophage and passes the phage-
Francis Crick and Sydney Brenner used E. coli genetics to establish the triplet nature of the genetic code in 1961. By 1966, the genetic code had been cracked by Marshall Nirenberg, Heinrich Matthaei, Gobind Khorana, and their colleagues, using synthetic oligonucleotides and E. coli extracts (see Chapter 17). A completely new era of gene regulation research was opened up by the studies of François Jacob and Jacques Monod on the detailed workings of the lac operon (see Chapter 20 and the How We Know section at the end of Chapter 5). The identification of plasmids in E. coli, and methods to handle them, ushered in the era of recombinant DNA technology. Plasmids continue to be wonderful biotech tools, valuable beyond compare (see Chapter 7).
Life Cycle
Like most bacteria, E. coli reproduces by binary fission (Figure A-1). As the cell grows, it senses when it has reached the correct size to begin replication. The duplicate chromosomes partition to opposite poles of the cell. Cytokinesis splits the cell in half to produce two daughter cells, each containing an identical chromosome. The entire process requires only about 20 minutes at 37°C, the average mammalian body temperature. Thus, a large, visible colony of millions of identical bacteria form within only 24 hours on a Petri plate of rich medium. Huge quantities of E. coli can also be grown in liquid culture within a day.
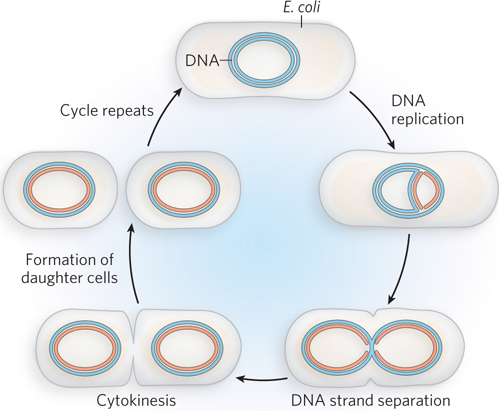
A-
Genetic Techniques
For a complete description of many of these techniques, see Chapter 7.
Mutagenesis Mutation frequency in E. coli can be enhanced by chemicals or UV light. Mutant cells can be selected by phenotype or screened by replica plating on different media.
Plasmids E. coli has several natural plasmids that can harbor foreign DNA in the cell. Researchers use these plasmids for protein expression, as shuttle vectors, or for building and sequencing large genomes.
Introduction of DNA E. coli can be made to accept foreign DNA by chemical transformation or electroporation. Cells transformed with plasmid DNA containing an antibiotic-
Gene Knockouts Plasmids without a replication origin but containing an antibiotic-
Complementation This technique is often used to prove that a mutation is in a particular gene by adding a wild-
Recombinant Protein Expression Rapid and inexpensive growth of E. coli makes it the most widely used vehicle for the expression and purification of foreign proteins. Foreign proteins are typically expressed from genes located on high-
E. coli as a Model Organism Today
Multiprotein Machines Biochemical and structural studies are now illuminating the once unknown atomic details of structures and processes of information transfer. These include the structure and function of the DNA replication apparatus and the DNA repair and recombination proteins, regulation of RNA polymerase, and the structure and chemistry of the ribosome. These multiprotein structures and processes occur in bacteria and eukaryotes alike, but they are streamlined in the relatively simple E. coli cell.
Cytologic Studies Processes can be visualized in living E. coli cells through the use of fluorescent fusion proteins. For example, recent studies of this sort have allowed detection of the proteins involved in chromosomal replication and their movements along the chromosome during replication.
Recombinant DNA Technology E. coli holds the central position in recombinant DNA technology. Its plasmids are widely used as shuttle vectors to amplify and maintain the DNA libraries of other organisms. E. coli is also still one of the most widely used vehicles for the expression and purification of foreign proteins.
Systems Biology Its relatively small genome makes E. coli an attractive system in which to pioneer approaches to systems biology. DNA chips have been constructed to examine the response of every transcript to a wide variety of conditions. The functions of about 35% of E. coli genes are still unknown; a genome-
A-