Chapter Introduction
The Dynamic
Genome: Transposable
Elements
CHAPTER
15
LEARNING OUTCOMES
After completing this chapter, you will be able to
Describe how transposable elements were first discovered genetically in maize and then first isolated molecularly from E. coli.
Describe how transposable elements participate in the spread of antibiotic-
resistant bacteria. Compare and contrast the two major mechanisms used by elements to transpose.
Provide reasons to explain how the human species thrives, with more than 50 percent of our genome derived from transposable elements.
Describe the mechanisms used by host genomes to repress the spread of some transposable elements.
Describe the strategies used by transposable elements to avoid host repression mechanisms.
A boy is born with a disease that makes his immune system ineffective. Diagnostic testing determines that he has a recessive genetic disorder called SCID (severe combined immunodeficiency disease), more commonly known as bubble-

In the past two decades, techniques have been developed that offer the possibility of a different kind of transplantation therapy—
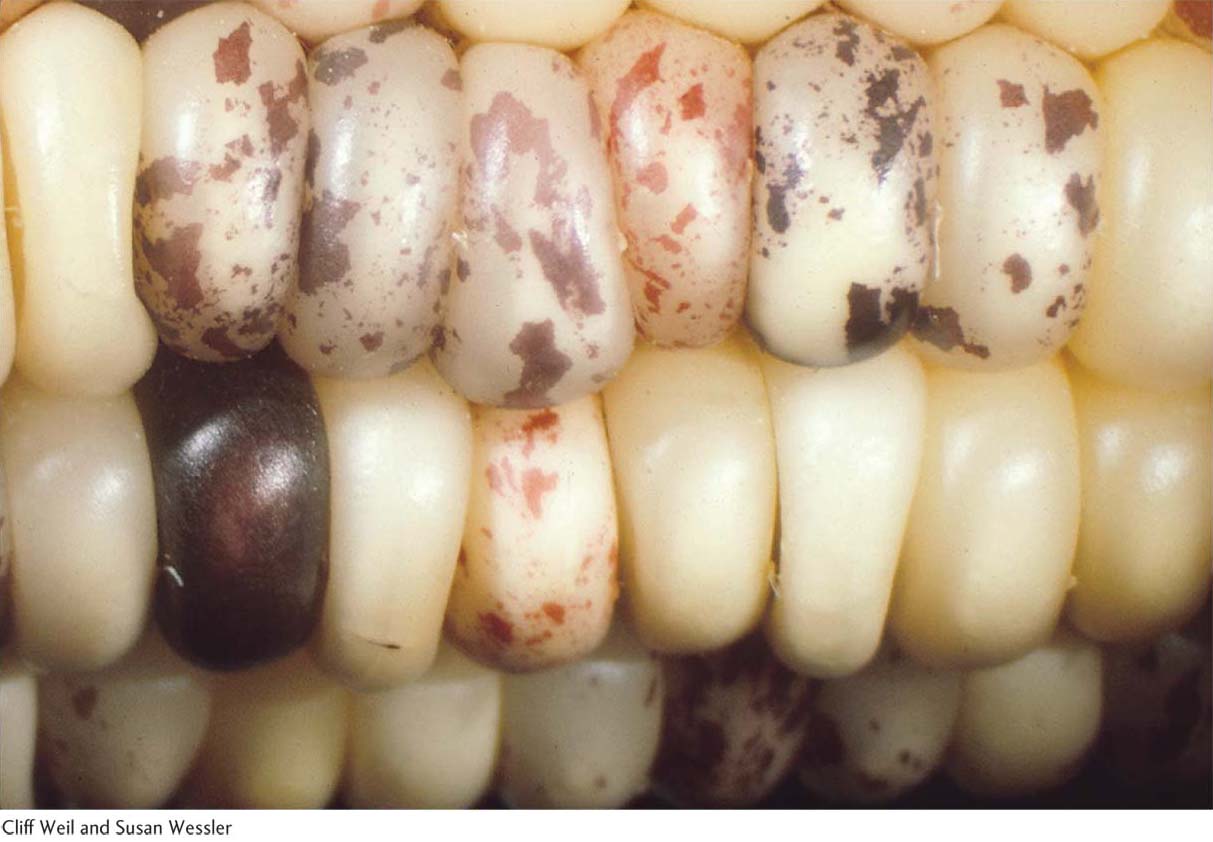
Starting in the 1930s, genetic studies of maize yielded results that greatly upset the classical genetic picture of genes residing only at fixed loci on chromosomes. The research literature began to carry reports suggesting that certain genetic elements present in the main chromosomes can somehow move from one location to another. These findings were viewed with skepticism for many years, but it is now clear that such mobile elements are widespread in nature.
A variety of colorful names (some of which help to describe their respective properties) have been applied to these genetic elements: controlling elements, jumping genes, mobile genes, mobile elements, and transposons. Here we use the terms transposable elements and mobile elements, which embrace the entire family of types. Transposable elements can move to new positions within the same chromosome or even to a different chromosome. They have been detected genetically in model organisms such as E. coli, maize, yeast, C. elegans, and Drosophila through the mutations that they produce when they insert into and inactivate genes.
DNA sequencing of genomes from a variety of microbes, plants, and animals indicates that transposable elements exist in virtually all organisms. Surprisingly, they are by far the largest component of the human genome, accounting for almost 50 percent of our chromosomes. Despite their abundance, the normal genetic role of these elements is not known with certainty.
In their studies, scientists are able to exploit the ability of transposable elements to insert into new sites in the genome. Transposable elements engineered in the test tube are valuable tools, both in prokaryotes and in eukaryotes, for genetic mapping, creating mutants, cloning genes, and even producing transgenic organisms.
Let us reconstruct some of the steps in the evolution of our present understanding of transposable elements. In doing so, we will uncover the principles guiding these fascinating genetic units.