Nuclear Bodies Are Functionally Specialized Nuclear Domains
High-resolution visualization of plant- and animal-cell nuclei by electron microscopy and by staining with fluorescently labeled antibodies has revealed domains in nuclei in addition to chromosome territories and nucleoli. These specialized nuclear domains, called nuclear bodies, are not surrounded by membranes. Nonetheless, they are regions of high concentrations of specific proteins and RNAs that form distinct, often roughly spherical structures within the nucleus (Figure 10-46). The most prominent nuclear bodies are nucleoli, the sites of ribosomal subunit synthesis and assembly discussed earlier. Several other types of nuclear bodies have also been described in structural studies.
[Part (a) reprinted by permission from Macmillan Publishers Ltd., from Gall, J. G., “The centennial of the Cajal body,” Nat. Rev. Mol. Cell Biol., 2003, 4(12):975–980; permission conveyed through Copyright Clearance Center, Inc. Part (b) republished with permission of Elsevier, from Handwerger, K. E. and Gall, J. G., “Subnuclear organelles: new insights into form and function,” Trends Cell Biol. 2006, 16(1):19–26; permission conveyed through Copyright Clearance Center, Inc. Part (c) from Fox, A. H., and Lamond, A. I., “Paraspeckles,” Cold Spring Harb. Perspect. Biol., 2010, 2(7):a000687. Part (d) republished with permission of American Society for Microbiology, from Pennella, M. A., et al., “Adenovirus E1B 55-kilodalton protein is a p53-SUMO1 E3 ligase that represses p53 and stimulates its nuclear export through interactions with promyelocytic leukemia nuclear bodies,” J. Virol., 2010, 84(23):12210–25.]
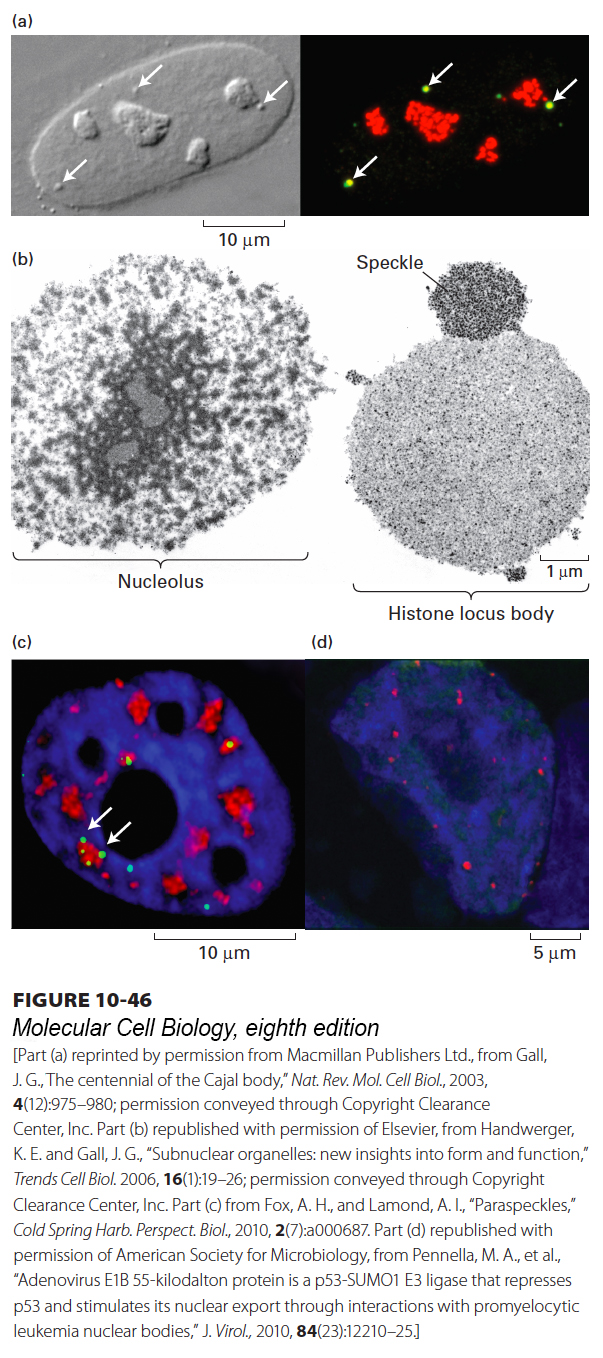
FIGURE 10-46Examples of nuclear bodies. (a) Cajal bodies and nucleoli in a HeLa cell nucleus. A DIC microscopic image (left) shows four nucleoli and three Cajal bodies (arrowheads). When the same nucleus is immunostained (right) with antibodies against coilin (green) and fibrillarin (red), the three Cajal bodies appear yellow because they stain with both antibodies. The nucleoli stain only for fibrillarin, which is the methyl transferase for 2′-O-methylation of rRNA in the nucleoli and snRNAs in the Cajal bodies. (b) Transmission electron micrograph of nuclear bodies in a single Xenopus oocyte nucleus. Only a portion of the extraordinarily large oocyte nucleus is shown. Histone locus bodies are sites of histone mRNA transcription and processing. They are larger in oocytes, which produce prodigious amounts of histone mRNAs, than in most vertebrate cells. A speckle is a region of concentrated splicing factors. (c) HeLa cell stained with DAPI (blue); antibody to SC35 (red), a splicing factor stored in speckles; and antibody to PSPC1, a protein found in nuclear bodies called paraspeckles (white arrows) because they are most often observed close to speckles. (d) PML nuclear bodies in the nucleus of an H1299 cell (a lung carcinoma cell line). DNA was stained with DAPI (blue) and PML nuclear bodies were immunostained with antibody to the major protein in these bodies, PML.
[Part (a) reprinted by permission from Macmillan Publishers Ltd., from Gall, J. G., “The centennial of the Cajal body,” Nat. Rev. Mol. Cell Biol., 2003, 4(12):975–980; permission conveyed through Copyright Clearance Center, Inc. Part (b) republished with permission of Elsevier, from Handwerger, K. E. and Gall, J. G., “Subnuclear organelles: new insights into form and function,” Trends Cell Biol. 2006, 16(1):19–26; permission conveyed through Copyright Clearance Center, Inc. Part (c) from Fox, A. H., and Lamond, A. I., “Paraspeckles,” Cold Spring Harb. Perspect. Biol., 2010, 2(7):a000687. Part (d) republished with permission of American Society for Microbiology, from Pennella, M. A., et al., “Adenovirus E1B 55-kilodalton protein is a p53-SUMO1 E3 ligase that represses p53 and stimulates its nuclear export through interactions with promyelocytic leukemia nuclear bodies,” J. Virol., 2010, 84(23):12210–25.]
Experiments with fluorescently labeled nuclear proteins have shown that the nucleus is a highly dynamic environment, in which proteins diffuse rapidly through the nucleoplasm. Proteins associated with nuclear bodies are often also observed at lower concentrations in the nucleoplasm outside the nuclear bodies, and fluorescence studies indicate that they diffuse into and out of the nuclear bodies. Based on measurements of molecular mobility in live cells, nuclear bodies can be mathematically modeled as the expected steady state for specific diffusing proteins that interact with sufficient affinity to form self-organized regions of high concentrations, but with low enough affinity for one another to be able to diffuse into and out of these structures. In electron micrographs, these structures appear as a heterogeneous, spongelike network of interacting components. We discuss a few examples of nuclear bodies here.
Cajal Bodies Cajal bodies are 0.2–1-µm spherical structures that have been observed in large nuclei for more than a century (Figure 10-46a). Current research indicates that, like nucleoli, Cajal bodies are centers of RNP-complex assembly for spliceosomal snRNPs and other RNPs. Like rRNAs, snRNAs undergo specific post-transcriptional modifications, such as the conversion of specific uridine residues to pseudouridine and the addition of methyl groups to the 2′-hydroxyl groups of specific riboses, that are important for the proper assembly and function of snRNPs in pre-mRNA splicing. These modifications occur in Cajal bodies, where they are directed by a class of snoRNA-like guide RNA molecules called scaRNAs (small Cajal body–associated RNAs). There is also evidence that Cajal bodies are sites of reassembly of the U4/U6/U5 tri-snRNP complex from the free U4, U5, and U6 snRNPs released during the removal of introns from mRNAs (see Figure 10-11).
Page 469
Histone Locus Bodies Histone locus bodies (Figure 10-46b) are sites of histone mRNA synthesis. They contain a high concentration of the U7 snRNP involved in the specialized 3′-end processing of the major histone mRNAs, which do not have a poly(A) tail.
Nuclear Speckles Nuclear speckles have been observed, using fluorescently labeled antibodies to snRNP proteins and other proteins involved in pre-mRNA splicing, as approximately 25–50 irregular, amorphous structures 0.5–2 µm in diameter distributed through the nucleoplasm of a vertebrate cell (Figure 10-46c). Because speckles are not located at sites of co-transcriptional pre-mRNA splicing, which are associated closely with chromatin, they are thought to be storage regions for snRNPs and proteins involved in pre-mRNA splicing that are released into the nucleoplasm when required.
Nuclear Paraspeckles Paraspeckles are composed of RNPs formed by the interaction between a long nonprotein-coding RNA species (lncRNA), NEAT1, and members of the DBHS (Drosophila Behavior Human Splicing) family of proteins, P54NRB/NONO, PSPC1, and PSF/SFPQ. Paraspeckles are critical to the control of gene expression through the nuclear retention of RNA containing double-stranded RNA regions that have been subjected to adenosine-to-inosine editing. In this way, they may function in the poorly understood mRNA quality-control mechanisms that operate in the nucleus.
Promyelocytic Leukemia (PML) Nuclear Bodies The PML gene was originally discovered when chromosomal translocations within it were observed in the leukemic cells of patients with a rare disease called promyelocytic leukemia (PML). When antibodies specific for the PML protein were used in immunofluorescence microscopy studies of mammalian cells, the protein was found to localize to 10–30 roughly spherical regions 0.3–1 µm in diameter in the cell nuclei. Multiple functions have been proposed for these PML nuclear bodies, but a consensus is emerging that they function as sites for the assembly and modification of protein complexes involved in DNA repair and the induction of apoptosis. For example, the important p53 tumor suppressor protein appears to be post-translationally modified by phosphorylation and acetylation in PML nuclear bodies in response to DNA damage, increasing its ability to activate the expression of genes whose products mitigate that damage. PML nuclear bodies are also required for cellular defenses against DNA viruses that are induced by interferons, proteins secreted by virus-infected cells and T-lymphocytes involved in the immune response (see Chapter 23).
PML nuclear bodies are also sites of post-translational modification of proteins through the addition of a small, ubiquitin-like protein called SUMO1 (small ubiquitin-like moiety-1), which can control the activity and subcellular localization of the modified protein. Many transcriptional activators are inhibited when they are sumoylated, and mutation of their site of sumoylation increases their activity in stimulating transcription. These observations indicate that PML nuclear bodies are involved in a mechanism of transcriptional repression that remains to be thoroughly understood.
Nucleolar Functions in Addition to Ribosomal Subunit Synthesis The first nuclear bodies to be observed, the nucleoli, have specialized regions of substructure (see Figure 10-46b) that are dedicated to functions other than ribosome biogenesis. There is evidence that the signal recognition particles involved in protein secretion and ER membrane insertion (see Chapter 13) are assembled in nucleoli and then exported to the cytoplasm, where their final maturation takes place. The Cdc14 protein phosphatase that regulates processes in the final stages of mitosis in yeast is sequestered in nucleoli until chromosomes have been properly segregated into the bud (see Chapter 19). In addition, a tumor suppressor protein called ARF, which is involved in the regulation of the protein encoded by the most frequently mutated gene in human cancers, the p53 gene, is sequestered in nucleoli and released in response to DNA damage (see Chapter 24). Furthermore, heterochromatin often forms on the surfaces of nucleoli (see Figure 8-28), suggesting that proteins associated with nucleoli participate in the formation of this transcription-repressing chromatin structure.