V-Class H+ ATPases Maintain the Acidity of Lysosomes and Vacuoles
All V-class ATPases transport only H+ ions. These proton pumps, present in the membranes of lysosomes, endosomes, and plant vacuoles, function to acidify the lumina of these organelles. The pH of the lysosomal lumen can be measured precisely in live cells by use of particles labeled with a pH-sensitive fluorescent dye. When these particles are added to the extracellular fluid, the cells engulf and internalize them (phagocytosis; see Figure 1-18 and Chapter 17), ultimately transporting them into lysosomes. The lysosomal pH can be calculated from the spectrum of the fluorescence emitted. The DNA encoding a naturally fluorescent protein whose fluorescence depends on the pH can be modified (by adding DNA segments encoding “signal sequences,” detailed in Chapters 13 and 14) such that the protein is targeted to the lysosome lumen; fluorescence measurements can then be used to determine the pH in the organelle lumen. Maintenance of the hundredfold or more proton gradient between the lysosomal lumen (pH ~4.5–5.0) and the cytosol (pH ~7.0) depends on a V-class ATPase and thus on ATP production by the cell. The low lysosomal pH is necessary for optimal function of the many proteases, nucleases, and other hydrolytic enzymes in the lumen; on the other hand, a cytosolic pH of 5 would disrupt the functions of many proteins optimized to act at pH 7 and lead to death of the cell.
Page 490
Pumping of relatively few protons is required to acidify an intracellular vesicle. To understand why, recall that a solution of pH 4 has a H+ ion concentration of 10−4 moles per liter, or 10−7 moles of H+ ions per milliliter. There are 6.02 × 1023 atoms of H per mole (Avogadro’s number), so a milliliter of a pH 4 solution contains 6.02 × 1016 H+ ions. Thus at pH 4, a primary spherical lysosome with a volume of 4.18 × 10−15 ml (diameter of 0.2 µm) would contain just 252 protons. At pH 7, the same organelle would have an average of only 0.2 protons in its lumen, and thus pumping of only about 250 protons would be necessary for lysosome acidification.
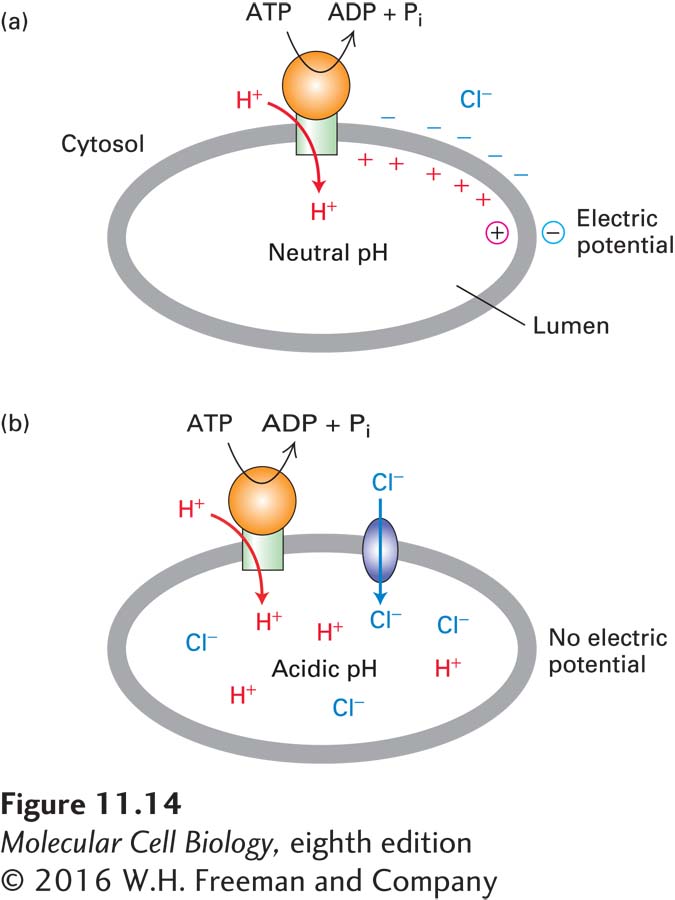
FIGURE 11-14Effect of V-class proton pumps on H+ concentration gradients and electric potential gradients across cellular membranes. (a) If an intracellular organelle contains only V-class pumps, proton pumping generates an electric potential across the membrane (the cytosolic face becomes negative and the luminal face positive), but no significant change in the intraluminal pH. (b) If the organelle membrane also contains Cl− channels, anions passively follow the pumped protons, resulting in an accumulation of H+ and Cl− ions in the lumen (low luminal pH) but no electric potential across the membrane.
By themselves, V-class proton pumps cannot acidify the lumen of an organelle (or the extracellular space) because these pumps are electrogenic; that is, a net movement of electric charge occurs during transport. Pumping of just a few protons causes a buildup of positively charged H+ ions on the exoplasmic (inside) face of the organelle membrane. For each H+ pumped across, a negative ion (e.g., OH− or Cl−) will be “left behind” on the cytosolic face, causing a buildup of negatively charged ions there. These oppositely charged ions attract each other on opposite faces of the membrane, generating a charge separation, or electric potential, across the membrane. The lysosome membrane thus functions as a capacitor in an electric circuit, storing opposing charges (anions and cations) on opposite sides of a barrier impermeable to the movement of charged particles.
As more and more protons are pumped and build up excess positive charge on the exoplasmic face, the energy required to move additional protons against this rising electric potential gradient increases dramatically and prevents pumping of additional protons long before a significant transmembrane H+ concentration gradient is established (Figure 11-14a). In fact, this is the way that P-class proton pumps generate a cytosol-negative potential across plant and yeast plasma membranes.
In order for an organelle lumen or an extracellular space (e.g., the lumen of the stomach) to become acidic, movement of protons must be accompanied either by (1) movement of an equal number of anions (e.g., Cl−) in the same direction or by (2) movement of equal numbers of a different cation in the opposite direction. The first process occurs in lysosomes and plant vacuoles, whose membranes contain V-class H+ ATPases and anion channels through which accompanying Cl− ions move (Figure 11-14b). The second process occurs in the lining of the stomach, which contains a P-class H+/K+ ATPase that is not electrogenic and pumps one H+ outward and one K+ inward. Operation of this pump is discussed later in the chapter.
Page 491
The V-class proton pumps in lysosomal and vacuolar membranes have been solubilized, purified, and incorporated into liposomes. As shown in Figure 11-9, these pumps contain two discrete domains, a cytosolic hydrophilic domain (V1) and a transmembrane domain (V0), with multiple subunits forming each domain. Binding and hydrolysis of ATP by the B subunits in V1 provide the energy for the pumping of H+ ions through the proton-conducting channel formed by the c and a subunits in V0. Unlike P-class ion pumps, V-class proton pumps are not phosphorylated and dephosphorylated during proton transport.