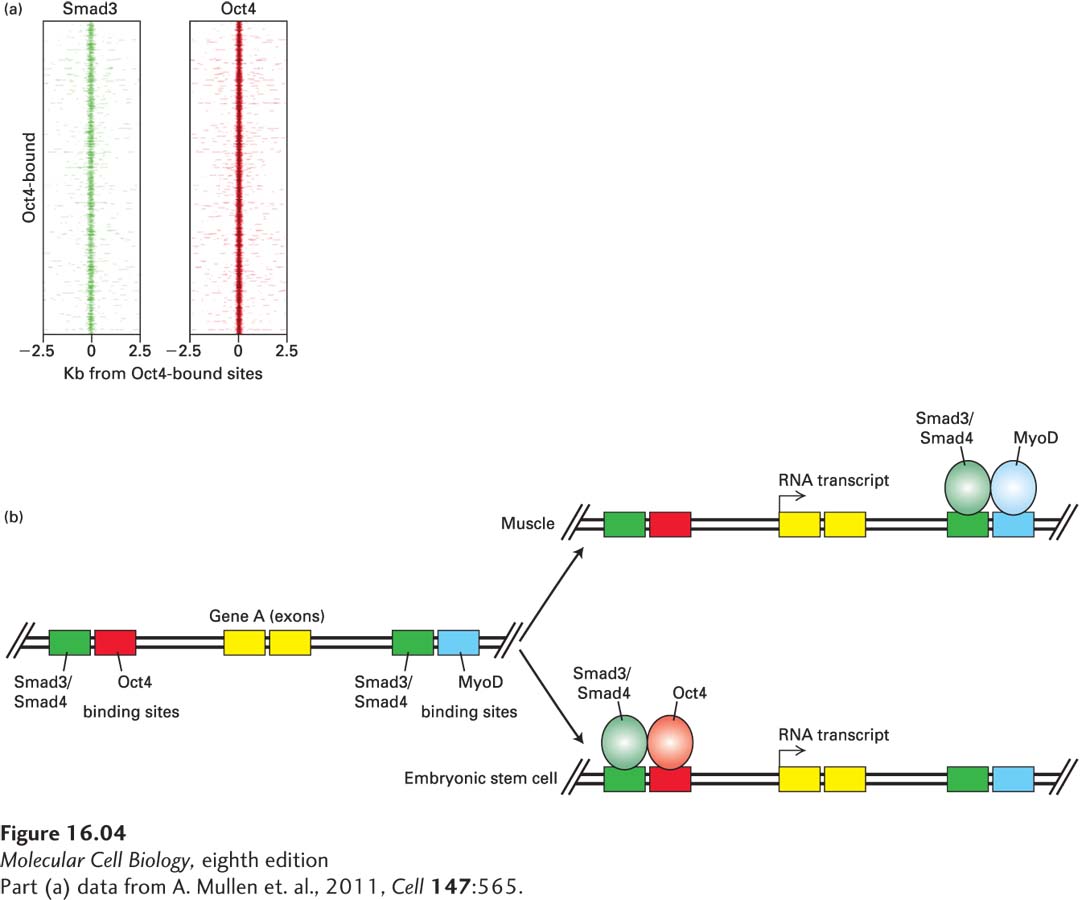
The Smad3/Smad4 Complex Activates Expression of Different Genes in Different Cell Types
Virtually all mammalian cells secrete at least one TGF-β isoform, and most have TGF-β receptors on their surface. However, the cellular responses induced by TGF-β vary among cell types. In epithelial cells and fibroblasts, for example, TGF-β induces expression of extracellular-matrix proteins (e.g., fibronectins and collagens; see Chapter 20). It also induces expression of proteins that inhibit serum proteases, which otherwise would degrade these extracellular-matrix proteins. This inhibition stabilizes the matrix, allowing cells to form stable tissues. The protease-inhibitory proteins include plasminogen activator inhibitor 1 (PAI-1). Transcription of the PAI-1 gene requires formation of a complex of the transcription factor TFE3 with the R-Smad/co-Smad (Smad3/Smad4) complex and binding of all these proteins to specific sequences within the regulatory region of the PAI-1 gene (Figure 16-3, bottom). By partnering with other transcription factors expressed in fibroblasts, the R-Smad/co-Smad complex promotes expression of genes encoding other proteins such as p15INK4B, which arrests the cell cycle at the G1 stage and thus blocks cell proliferation (see Chapter 19).
More generally, in order for the Smad3/Smad4 complex to bind to a DNA regulatory region and activate a given gene, the DNA binding segment must be in an active “open” chromatin conformation. Equally important, binding of the Smad3/Smad4 complex requires binding of other transcription factors at adjacent sites in the DNA; often these transcription factors are master transcription factors that determine the identity of a cell during its development. For example, we will learn in Chapter 21 that the transcription factor Oct4 forms a complex with two other transcription factors, Sox2 and Nanog, that are essential for keeping embryonic stem (ES) cells in an undifferentiated state, and that expression of these three master transcription factors, together with one or two other transcription factors, is sufficient to reprogram a differentiated cell (e.g., a cell of the immune system) into an ES cell. TGF–β activation of the Smad3/Smad4 complex is also required to maintain ES cells in an undifferentiated state.
The chromatin immunoprecipitation–DNA sequence experiment summarized in Figure 16-4a shows that in ES cells, the Smad3/Smad4 complex binds to DNA at sites adjacent to those occupied by Oct4; similar experiments showed that Sox2 and Nanog are also bound at most of these sites. In contrast, MyoD and the related transcription factor Myf5 are the master transcription factors in development of striated muscle, and Oct4, Sox2, and Nanog are not expressed; in muscle cells, the Smad3/Smad4 complex binds to gene regulatory sites adjacent to those occupied by MyoD, which are very different DNA sites from those that bind the Smad3/Smad4 complex in ES cells. About 13 percent of the genes in muscle whose regulatory regions are bound by the Smad3/Smad4 complex in muscle cells are also bound by the Smad3/Smad4 complex in ES cells, but detailed analyses of these binding sites showed that the Smad3/Smad4 complex regulates the same gene in these different cell types by binding to different regulatory DNA sequences, most presumably enhancer regions, which are occupied by different cell-specific master transcription factors (Figure 16-4b). As we noted earlier, other types of transcription factors activated by other types of cell-surface receptors also activate or repress different sets of genes in different types of cells. We will see in this chapter that those transcription factors regulate those genes in the same manner as the Smads activated by the TGF-β receptors.
[Part (a) data from A. Mullen et al., 2011, Cell147:565.]
EXPERIMENTAL FIGURE 16-4The Smad3/Smad4 complex binds to DNA at sites adjacent to those occupied by cell-specific master transcription factors. (a) In human embryonic stem cells (ES cells), the Smad3/Smad4 complex binds at sites near those occupied by the ES master transcription factor Oct4. Chromatin immunoprecipitation studies were performed using antibodies specific for Smad3 or Oct4 and the DNA sequences bound to these factors were determined; this process is often abbreviated “ChIP-Seq.” Plotted on the y axis are the top 1000 sites bound by Oct4; the intensity of the red color is proportional to the amount of DNA at this site bound by Oct4 within 2500 bases upstream (−) or downstream (+) of this binding site. The left panel shows binding of Smad3 to DNA sequences within 2500 bases of these 1000 Oct4 sites; it is apparent that most of the Oct4 sites have Smad3 bound near them. Similar plots centered around the top 1000 Smad3 sites showed that over 80 percent of the sites with bound Smad3 also had a bound Oct4. (b) Smad3 can regulate the same gene in different cell types by binding to regulatory DNA sequences occupied by different cell-specific master transcription factors. In this hypothetical scenario, the Smad3/Smad4 complex (green) activates the same gene in ES cells and in muscle cells. In ES cells, it co-occupies a regulatory site with Oct4 (red, binding to a site in DNA colored red), and in muscle, it co-occupies a different regulatory site with the muscle master transcriptional regulator, MyoD (blue).
[Part (a) data from A. Mullen et al., 2011, Cell147:565.]
Page 725
Loss of TGF-β signaling plays a key role in the early development of many cancers. Many human tumors contain inactivating mutations in either TGF-β receptors or Smad proteins and thus are resistant to growth inhibition by TGF-β (see Figure 24-24). Most human pancreatic cancers, for instance, contain a deletion in the gene encoding Smad4 and thus cannot induce cell cycle inhibitors in response to TGF-β. In fact, Smad4 was originally called DPC (deleted in pancreatic cancer). Retinoblastoma, colon and gastric cancer, hepatoma, and some T- and B-cell malignancies are also unresponsive to TGF-β growth inhibition. This loss of responsiveness correlates with loss of TGF-β RI or RII; responsiveness to TGF-β can be restored by recombinant expression of the “missing” protein. Mutations in Smad2 also commonly occur in several types of human tumors.