Eukaryotic Cells Contain a Large Number of Internal Membrane Structures
We noted earlier that, unlike prokaryotic cells, most eukaryotic cells contain extensive internal membranes that enclose specific subcellular compartments, termed organelles. Here we review the organelles and their functions.
Page 15
Page 16
Endoplasmic Reticulum and Golgi Complex Generally the largest membrane in a eukaryotic cell encloses the organelle termed the endoplasmic reticulum (ER)—an extensive network of closed, flattened membrane-
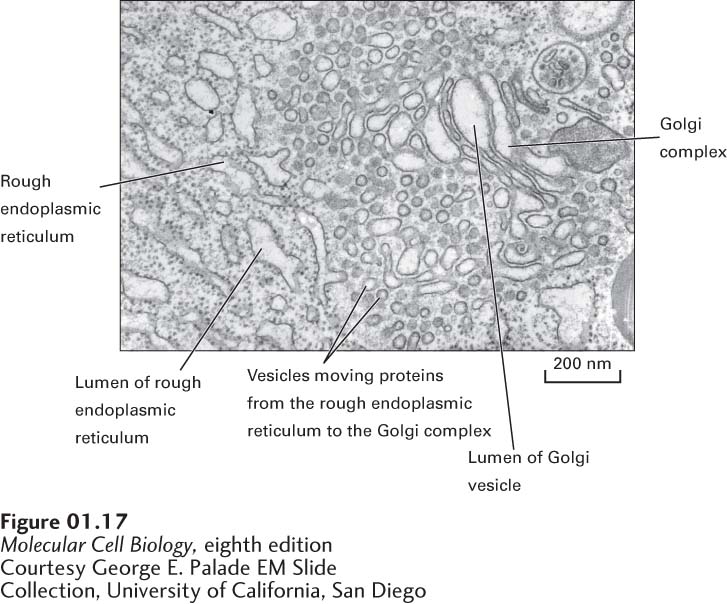
In contrast, the cytosolic side of the rough endoplasmic reticulum is studded with ribosomes; these ribosomes synthesize certain membrane and organelle proteins and virtually all proteins that are to be secreted from the cell (see Chapter 13). As a growing polypeptide emerges from a ribosome, it passes through the rough ER membrane with the help of specific transport proteins that are embedded in the membrane. Newly made membrane proteins remain associated with the rough ER membrane, and proteins to be secreted accumulate in the lumen, the aqueous interior of the organelle. Several minutes after proteins are synthesized in the rough ER, most of them leave the organelle within small membrane-
After proteins to be secreted and membrane proteins are modified in the Golgi complex, they are transported out of the complex by a second set of vesicles, which bud from one side of the Golgi complex. Some vesicles carry membrane proteins destined for the plasma membrane or soluble proteins to be released from the cell into the extracellular space; others carry soluble or membrane proteins to lysosomes or other organelles. How intracellular transport vesicles “know” with which membranes to fuse and where to deliver their contents is also discussed in Chapter 14.
EndosomesAlthough transport proteins in the plasma membrane mediate the movement of ions and small molecules into the cell across the lipid bilayer, proteins and some other soluble macromolecules in the extracellular milieu are internalized by endocytosis. In this process, a segment of the plasma membrane invaginates into a coated pit, whose cytosolic face is lined by a specific set of proteins that cause vesicles to form. The pit pinches from the membrane into a small membrane-
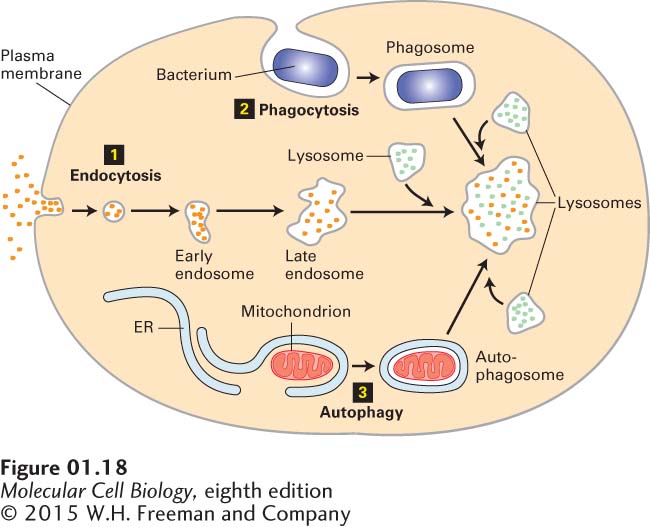
Lysosomes Lysosomes provide an excellent example of the ability of intracellular membranes to form closed compartments in which the composition of the lumen (the aqueous interior of the compartment) differs substantially from that of the surrounding cytosol. Found exclusively in animal cells, lysosomes are responsible for degrading many components that have become obsolete for the cell or organism. The process by which an aged organelle is degraded in a lysosome is called autophagy (“eating oneself”). Materials taken into a cell by endocytosis or phagocytosis may also be degraded in lysosomes (see Figure 1-18). In phagocytosis, large, insoluble particles (e.g., bacteria) are enveloped by the plasma membrane and internalized.
Page 17
Lysosomes contain a group of enzymes that degrade polymers into their monomeric subunits. For example, nucleases degrade RNA and DNA into their mononucleotide building blocks; proteases degrade a variety of proteins and peptides; phosphatases remove phosphate groups from mononucleotides, phospholipids, and other compounds; still other enzymes degrade complex polysaccharides and glycolipids into smaller units. All of these lysosomal enzymes, collectively termed acid hydrolases, work most efficiently at acidic pH values. The acidic pH helps to denature proteins, making them accessible to the action of the lysosomal hydrolases. These enzymes are less active at the neutral pH of cells and most extracellular fluids. Thus if a lysosome releases its enzymes into the cytosol, where the pH is between 7.0 and 7.3, they cause little degradation of cytosolic components. Cytosolic and nuclear proteins generally are not degraded in lysosomes, but rather in proteasomes, large multiprotein complexes in the cytosol (see Chapter 3).
Peroxisomes All animal cells (except erythrocytes) and many plant and fungal cells contain peroxisomes, a class of roughly spherical organelles 0.2–
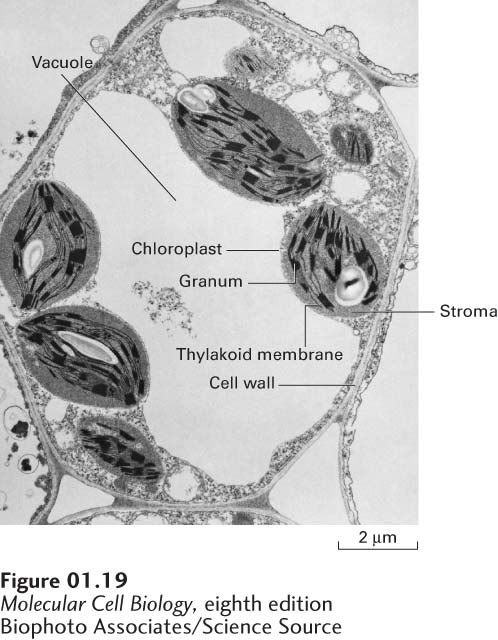
Plant Vacuoles Most plant cells contain at least one membrane-
Page 18