Gap Junctions Composed of Connexins Allow Small Molecules to Pass Directly Between the Cytosols of Adjacent Cells
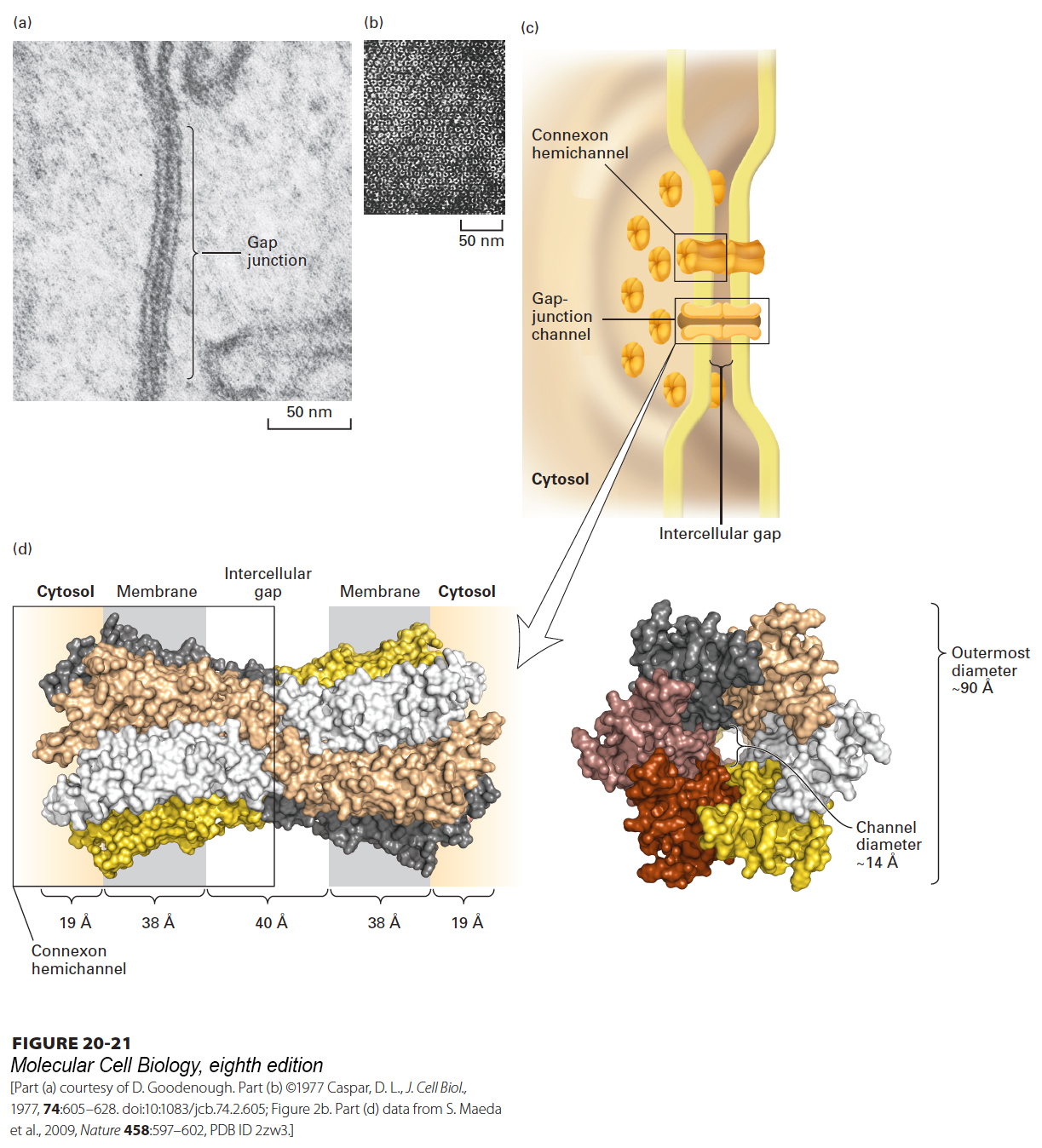
Early electron micrographs of tissues revealed sites of cell-cell contact with a characteristic intercellular gap (Figure 20-21a). This feature, which was found in virtually all animal cells that contact other cells, prompted early morphologists to call these regions gap junctions. In retrospect, the most important feature of these junctions is not the 2–4-nm gap itself, but a well-defined set of cylindrical particles that cross the gap and compose pores connecting the cytosols of adjacent cells (Figure 20-21b, c). As we will see later in this chapter, plant cells also assemble pores that connect the cytosols of adjacent cells, but those channels, called plasmodesmata, differ considerably in structure from gap junctions. Tunneling nanotubes are relatively recently discovered membrane-bound tubes that connect the cytoplasms of animal cells. Because they are more similar to plasmodesmata than to gap junctions, they will be discussed with plasmodesmata.
FIGURE 20-21Gap junctions. (a) In this thin section through a gap junction connecting two mouse liver cells, the two plasma membranes are closely associated for a distance of several hundred nanometers, separated by a “gap” of 2–3 nm. (b) Numerous roughly hexagonal particles are visible in this perpendicular view of the cytosolic face of a region of plasma membrane enriched in gap junctions. Each hexagonal particle aligns with a similar particle on an adjacent cell, forming a channel connecting the two cells. (c) Schematic model of a gap junction connecting two plasma membranes. Both membranes contain connexon hemichannels, cylinders of six dumbbell-shaped connexin molecules. Two connexons join in the gap between the cells to form a gap-junction channel, 1.4–2.0 nm in diameter, that connects the cytosols of the two cells. (d) Structure of recombinant human Cx26 gap junction as determined by x-ray crystallography (3.5-Å resolution). Left: Space-filling model of a side view of the complete structure of two attached connexons oriented as in part (c). Each of the six connexins that comprise a connexon has four transmembrane helices and is shown in a distinct color. The structures of the loops connecting the transmembrane helices are not well defined and not shown. Right: View from the cytosol perpendicular to the membrane bilayers, looking down on the connexon with its central pore. The diameter of the pore’s channel is ~14 Å, and it is lined by many polar/charged amino acids. See S. Nakagawa et al., 2010, Curr. Opin. Struct. Biol.20(4):423–430.
In many animal tissues, anywhere from a few to thousands of gap-junction particles cluster together in patches (e.g., along the lateral surfaces of epithelial cells; see Figure 20-11). When the plasma membrane is purified and then sheared into small fragments, some pieces mainly containing patches of gap junctions are generated. Owing to their relatively high protein content, these fragments have a higher density than the bulk of the plasma membrane and can be purified by equilibrium density-gradient centrifugation (see Figure 4-37). When these preparations are viewed perpendicular to the membrane, the gap junctions appear as arrays of hexagonal particles that enclose water-filled channels (see Figure 20-21b).
The effective pore size of gap junctions can be measured by injecting a cell with a fluorescent dye covalently linked to membrane bilayer–impermeable molecules of various sizes and observing with a fluorescence microscope whether the dye passes into neighboring cells. Gap junctions between mammalian cells permit the passage of molecules as large as 1.2 nm in diameter. In insects, these junctions are permeable to molecules as large as 2 nm in diameter. Generally speaking, molecules smaller than 1200 Da pass freely and those larger than 2000 Da do not pass; the passage of intermediate-sized molecules is variable and limited. Thus ions, many low-molecular-weight precursors of cellular macromolecules, products of intermediary metabolism, and small intracellular signaling molecules can pass from cell to cell through gap junctions.
In neural tissue, some neurons are connected by gap junctions through which ions pass rapidly, thereby allowing very rapid transmission of electrical signals. Impulse transmission through these connections, called electrical synapses, is almost a thousand times as rapid as at chemical synapses (see Chapter 22). Gap junctions are also present in many non-neural tissues, where they help to integrate the electrical and metabolic activities of many cells. In the heart, for instance, gap junctions rapidly pass ionic signals among cardiac muscle cells, which are tightly bound together via desmosomes. Thus gap junctions contribute to the electrically stimulated coordinate contraction of cardiac muscle cells during a heartbeat. As discussed in Chapter 15, some extracellular hormonal signals induce the production or release of small intracellular signaling molecules called second messengers (e.g., cyclic AMP, IP3, and Ca2+) that regulate cellular metabolism. Because many second messengers can be transferred between cells through gap junctions, hormonal stimulation of one cell has the potential of triggering a coordinated response by that cell as well as many of its neighbors. Such gap-junction-mediated signaling plays an important role, for example, in the secretion of digestive enzymes by the pancreas and in the coordinated muscular contractile waves (peristalsis) in the intestine. Another vivid example of gap-junction-mediated transport is the phenomenon of metabolic coupling, or metabolic cooperation, in which a cell transfers nutrients or intermediary metabolites to a neighboring cell that is itself unable to synthesize them. Gap junctions play critical roles in the development of egg precursors (oocytes) in the ovary by mediating the movement of both metabolites and signaling molecules, such as cyclic GMP, between an oocyte and its surrounding granulosa cells, as well as between neighboring granulosa cells.
A current model of the structure of the gap junction is shown in Figure 20-21c–d. Vertebrate gap junctions are composed of connexins, a family of structurally related transmembrane proteins with molecular weights between 26,000 and 60,000. Each vertebrate hexagonal particle consists of twelve noncovalently associated connexin molecules: six form a cylindrical hemichannel, called a connexon, in one plasma membrane that is joined to a connexon in the adjacent cell membrane, forming a continuous aqueous channel (diameter ~14 Å) between the cells. Each individual connexin molecule has four membrane-spanning α helices with a topology similar to that of claudin (see Figure 20-18), resulting in 24 transmembrane α helices in each connexon hemichannel.
Page 943
Page 944
A completely different family of proteins, the innexins, forms the gap junctions in invertebrates. A third family of innexin-like proteins, called pannexins, has been found in both vertebrates and invertebrates. Pannexins form hexamer hemichannels (pannexons) whose opening can be regulated by changes in membrane potential or mechanical stress. When open, pannexons permit direct exchange of small molecules (such as ATP) and ions between the intracellular and extracellular spaces. Pannexons are thought to play key roles in release of ATP from cells into the extracellular space. Extracellular ATP (as well as ADP and AMP) can function as an intercellular messenger or transmitter by binding to and activating the cell-surface purinergic receptors P1, P2X, or P2Y on target cells.
There are 21 different connexin genes in humans, and different sets of connexins are expressed in different cell types. The existence of this diversity, together with the generation of mutant mice with inactivating mutations in connexin genes, has highlighted the importance of connexins in a wide variety of cellular systems. Some cells express a single connexin that forms homotypic connexons. Most cells, however, express at least two connexins; these different proteins can assemble into heteromeric connexons, which in turn form heterotypic gap-junction channels. Diversity in channel composition leads to differences in channel permeability. For example, channels made from a 43-kDa connexin isoform, Cx43—the most ubiquitously expressed connexin—are more than a hundred times as permeable to ADP and ATP as those made from Cx32 (32 kDa).
The permeability of gap junctions is regulated by post-translational modification of connexins (e.g., phosphorylation) and is sensitive to changes in environmental conditions such as intracellular pH and Ca2+ concentration, membrane potential, and the intercellular potential between adjacent interconnected cells (“voltage gating”). The N-termini of connexins appear to be especially important in the gating mechanism. Thus, as is the case for many ion channels (see Chapter 11), the channel in some gap junctions can be either opened or closed. One example of the physiological regulation of gap junctions occurs during mammalian childbirth. The smooth muscle cells in the mammalian uterus must contract strongly and synchronously during labor to expel the fetus. To facilitate this coordinated activity, immediately before and during labor there is an approximately five- to tenfold increase in the amount of the major connexin in these cells, Cx43, and an increase in the number and size of gap junctions, which is reversed rapidly postpartum (following childbirth).
The assembly of connexins, their trafficking within cells, and the formation of functional gap junctions apparently depend on N-cadherin and its associated adapter proteins (e.g., α- and β-catenins, ZO-1, and ZO-2) as well as desmosomal proteins (plakoglobin, desmoplakin, and plakophilin-2). PDZ domains in ZO-1 and ZO-2 bind to the C-terminus of Cx43 and mediate its interaction with catenins and N-cadherin. The relevance of these relationships is particularly evident in the heart, which depends on gap junctions for rapid coordinated electrical coupling and on adjacent adherens junctions and desmosomes for mechanical coupling between cardiomyocytes to achieve the intercellular integration of electrical activity and movement required for normal cardiac function. It is noteworthy that ZO-1 serves as an adapter for adherens (see Figure 20-14), tight, and gap junctions, suggesting that this and other adapters can help integrate the formation and functions of these diverse junctions.
Mutations in connexin genes cause at least eight human diseases, including neurosensory deafness (Cx26 and Cx31), cataracts or heart malformations (Cx43, Cx46, and Cx50), and the X-linked form of Charcot-Marie-Tooth disease (Cx32), which is marked by progressive degeneration of peripheral nerves.