The Proteasome Is a Molecular Machine Used to Degrade Proteins
Proteasomes are very large protein-degrading molecular machines that influence many different cellular functions, including the cell cycle (see Chapter 19), transcription, DNA repair (see Chapter 5), programmed cell death, or apoptosis (see Chapter 21), recognition of and response to infection by foreign organisms (see Chapter 23), and removal of misfolded proteins. There are approximately 30,000 proteasomes in a typical mammalian cell.
Page 98
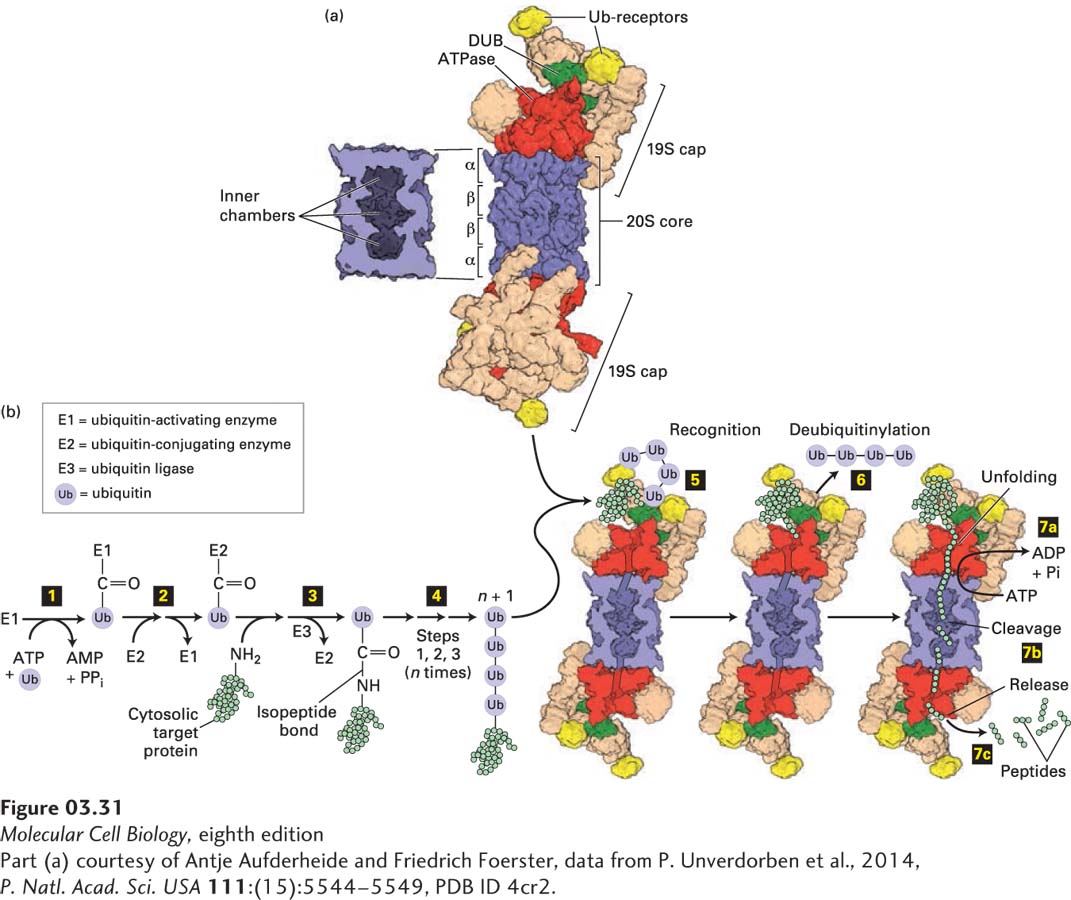
Proteasomes consist of roughly 60 protein subunits and have a mass of about 2.4 × 106 Da. Proteasomes have a cylindrical, barrel-like catalytic core (Figure 3-31a), called the 20S proteasome (where S is a Svedberg unit based on the sedimentation properties of the particle and is proportional to its size), which is approximately 14.8 nm tall and 11.3 nm in diameter. Bound to the ends of this core are either one or two 19S cap complexes that regulate the activity of the 20S catalytic core. When the core and one or two caps are combined, they are referred to collectively as the 26S complex, even though the two-cap-containing complex is larger (30S). A 19S cap has 19 protein subunits, six of which can hydrolyze ATP (i.e., they are AAA-type ATPases) to provide the energy needed to unfold protein substrates and selectively transfer them into the inner chamber of the proteasome’s catalytic core. Genetic studies in yeast have shown that cells cannot survive without functional proteasomes, thus demonstrating their importance. Furthermore, proper proteasomal activity is so important that cells will expend as much as 30 percent of the energy needed to synthesize a protein to degrade it in a proteasome.
[Part (a) courtesy of Antje Aufderheide and Friedrich Foerster, data from P. Unverdorben et al., 2014, P. Natl. Acad. Sci. USA111:(15):5544–5549, PDB ID 4cr2.]
FIGURE 3-31Ubiquitin- and proteasome-mediated proteolysis. (a) Right: The 26S proteasome has a cylindrical structure with a 19S cap at one or both ends of the 20S core particle. The nineteen different subunits of the 19S cap (shown in multiple colors) include six AAA-ATPase subunits (Rpt1–6, red), which assemble into a heterohexameric ring; two ubiquitin (Ub) receptors (Rpn10 and Rpn13, yellow); and a deubiquitinase enzyme (DUB, Rpn11, green), which forms a heterodimer with its evolutionarily related counterpart Rpn8. Moreover, the 19S cap contains scaffolding and other proteins (tan). The two 19S caps shown are facing in opposite directions relative to the plane of the page. The 20S core consists of four stacked heptameric rings (~110 Å diameter × 160 Å long), each containing either α (outer rings) or β (inner rings) subunits (blue). Left: Cutaway view of the 20S core, showing the inner chambers. Proteolysis occurs within the central inner chamber of the core formed by the β rings. (b) Proteins are targeted for proteasomal degradation by polyubiquitinylation. Enzyme E1 is activated by the ATP-dependent attachment of a ubiquitin (Ub) molecule (step 1) and then transfers this Ub molecule to a cysteine residue in E2 (step 2). Ubiquitin ligase (E3) transfers the bound Ub molecule on E2 to the side-chain –NH2 of a lysine residue in a target protein, forming an isopeptide bond (step 3). Additional Ub molecules are added to the Ub-modified target protein via isopeptide bonds to the previously added Ub by repeating steps 1 – 3, forming a polyubiquitin chain (step 4). The polyubiquitinylated target is recognized by Ub receptors in the proteasome’s 19S cap (step 5), and the Ub groups are removed by the deubiquitinase enzyme (step 6). In step 7 ATP hydrolysis enables the six protein (hexameric) ATPase subunits (red) to unfold the substrate and transfer the unfolded protein via a pore in the hexamer into the proteolysis chamber in the 20S core (step 7a), in some cases coordinately with step 6, and the protein is cleaved into short peptide digestion fragments (step 7b) that are then released (step 7c).
[Part (a) courtesy of Antje Aufderheide and Friedrich Foerster, data from P. Unverdorben et al., 2014, P. Natl. Acad. Sci. USA111:(15):5544–5549, PDB ID 4cr2.]
Page 99
The 20S proteasomal catalytic core comprises two inner rings of seven β subunits each, with three proteolytic active sites per ring facing toward the ~1.7-nm-diameter inner chamber formed by those rings, and two outer rings of seven α subunits each, which limit substrate access (see Figure 3-31a) via an entry channel that can be opened by the 19S cap. Proteasomes can degrade most proteins thoroughly because the three active sites in each β subunit ring can cleave peptide bonds at hydrophobic residues, acidic residues, or basic residues. Polypeptide substrates must enter the chamber via a regulated ~1.3-nm-diameter aperture at the center of the outer α subunit rings. In the 26S proteasome, the opening of the aperture, which is narrow and often allows the entry of only unfolded proteins, is controlled by ATPases in the 19S cap. These ATPases are responsible for unfolding protein substrates and translocating those unfolded polypeptides into the inner chamber of the catalytic core (Figure 3-31b, bottom right). The short peptide products of proteasomal digestion (2–24 residues long) exit the chamber and are further degraded rapidly by cytosolic peptidases, eventually being converted to individual (“free”) amino acids. One researcher has quipped that a proteasome is a “cellular chamber of doom” in which proteins suffer a “death by a thousand cuts.”
Inhibitors of proteasome function have proved to be exceptionally useful in the laboratory and the clinic. Small-molecule proteasome inhibitors, such as MG132, are used to block proteasomal degradation in the lab and to help evaluate the role of the proteasome and, as we shall see below, polyubiquitinylation in a wide variety of processes. Other small-molecule proteasome inhibitors have been used therapeutically. Because of the global importance of proteasome-mediated protein breakdown in cells, continuous, complete inhibition of proteasomes kills cells. However, partial proteasome inhibition for short intervals is widely used as an approach to cancer chemotherapy, especially to treat multiple myeloma, a cancer involving the abnormal proliferation of antibody (immunoglobulin)-producing cells. The myeloma cells produce abnormally high levels of potentially toxic, aberrant immunoglobulin polypeptide chains, which are degraded by proteasomes. Proteasome inhibition in these cancer cells leads to the buildup of toxic, misfolded immunoglobulin polypeptides within the cells, and thus to cell death. In addition, to survive and grow, myeloma cells require the robust activity of a regulatory protein called NF-κB (see Chapter 16) as well as other “pro-survival” and “pro-proliferation” proteins. In turn, NF-κB can function fully and promote survival and proliferation only when its inhibitor, I-κB, is disengaged and degraded by proteasomes (see Chapter 16). Partial inhibition of proteasomal activity by a small-molecule inhibitor drug results in increased levels of I-κB and, consequently, reduced NF-κB activity (that is, loss of its protective activity). The cancer cells subsequently undergo less proliferation and die by apoptosis. Thus, multiple myeloma cells are more sensitive to proteasome inhibitors than normal cells. Consequently, controlled administration of proteasome inhibitors, at levels that kill the cancer cells but not normal cells, has proved to be an effective therapy for multiple myeloma.