Noncovalent Binding of Calcium and GTP Are Widely Used as Allosteric Switches to Control Protein Activity
Unlike oxygen, which causes graded allosteric changes in the activity of hemoglobin, some other allosteric effectors act as switches, turning the activity of many different proteins on or off by binding to them noncovalently. Two important allosteric switches that we will encounter many times throughout this book, especially in the context of cell signaling pathways (see Chapters 15 and 16), are Ca2+ and GTP.
Ca2+/Calmodulin-Mediated Switching The concentration of Ca2+ that is free in the cytosol (not bound to molecules other than water) is kept very low (~10−7 M) by specialized membrane transport proteins that continually pump excess Ca2+ out of the cytosol (see Chapters 11 and 15). However, as we will learn in Chapters 11 and 15, the cytosolic Ca2+ concentration can increase tenfold to a hundredfold when Ca2+-permeable channels in the cell-surface membranes open and allow extracellular Ca2+ to flow into the cell. This rise in cytosolic Ca2+ is sensed by specialized Ca2+-binding proteins, which alter cellular behavior by turning the activities of other proteins on or off. The importance of extracellular Ca2+ for cell activity was first documented by S. Ringer in 1883, when he discovered that isolated rat hearts suspended in an NaCl solution made with “hard” (Ca2+-rich) London tap water contracted beautifully, whereas they beat poorly and stopped quickly if distilled, Ca2+-depleted, water was used.
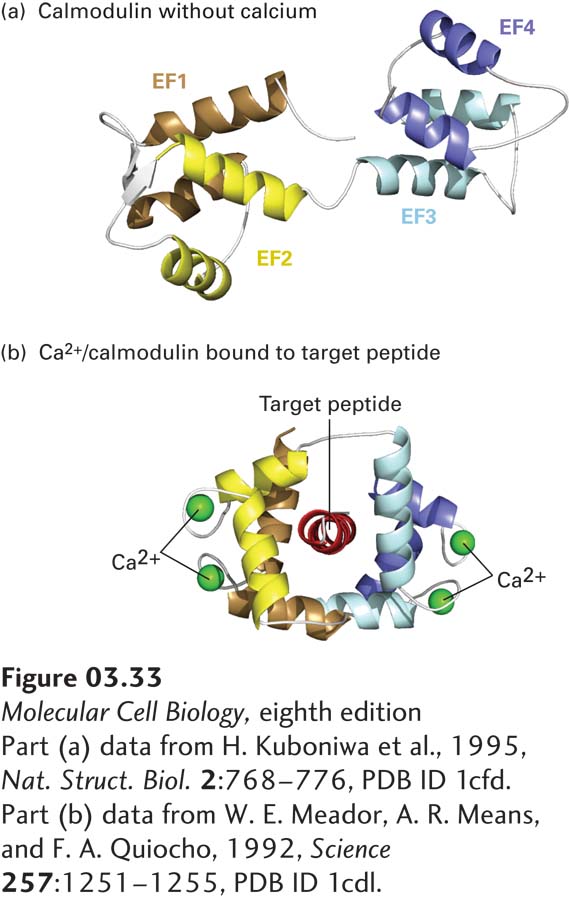
Many Ca2+-binding proteins bind Ca2+ using the EF hand/helix-loop-helix structural motif discussed earlier (see Figure 3-10b). A well-studied EF hand protein, calmodulin, is found in all eukaryotic cells, where it may exist as an individual monomeric protein or as a subunit of a multimeric protein. This dumbbell-shaped molecule contains four Ca2+-binding EF hands with Kds of about 10−6 M. The binding of Ca2+ to calmodulin causes a conformational change that permits Ca2+/calmodulin to bind to conserved sequences in various target proteins (Figure 3-33), thereby switching their activities on or off. Calmodulin and similar EF hand proteins thus function as switch proteins, acting in concert with changes in Ca2+ levels to modulate the activity of other proteins.
[Part (a) data from H. Kuboniwa et al., 1995, Nat. Struct. Biol.2:768–776, PDB ID 1cfd. Part (b) data from W. E. Meador, A. R. Means, and F. A. Quiocho, 1992, Science257:1251–1255, PDB ID 1cdl.]
FIGURE 3-33Conformational changes induced by Ca2+ binding to calmodulin. Calmodulin is a widely distributed cytosolic protein that contains four Ca2+-binding sites, one in each of its EF hand (helix-loop-helix) motifs (EF1-ER4, see also Figure 3-10). At cytosolic Ca2+ concentrations above about 5 × 10−7 M, binding of Ca2+ to calmodulin changes the protein’s conformation from the dumbbell-shaped, Ca2+-free form (a) to one in which hydrophobic side chains become more exposed to solvent. The resulting Ca2+/calmodulin complex can wrap around exposed helices (target peptides) with specialized sequences in various target proteins (b), thereby altering their activities.
[Part (a) data from H. Kuboniwa et al., 1995, Nat. Struct. Biol.2:768–776, PDB ID 1cfd. Part (b) data from W. E. Meador, A. R. Means, and F. A. Quiocho, 1992, Science257:1251–1255, PDB ID 1cdl.]
Page 102
Switching Mediated by Guanine Nucleotide–Binding Proteins Another group of intracellular switch proteins constitutes the GTPase superfamily. As the name suggests, these proteins are enzymes—GTPases—that can hydrolyze GTP (guanosine triphosphate) to GDP (guanosine diphosphate). They include the monomeric Ras protein (whose structure is shown in Figure 3-9, with bound GDP shown in blue) and the Gα subunit of the trimeric G proteins, both discussed at length in Chapters 15 and 16. Both Ras and Gα can bind to the plasma membrane, function in cell signaling, and play key roles in cell proliferation and differentiation. Other members of the GTPase superfamily function in protein synthesis, the transport of proteins between the nucleus and the cytoplasm, the formation of coated vesicles and their fusion with target membranes, and rearrangements of the actin cytoskeleton. Some GTPase proteins have a covalently attached lipid chain (see Figure 7-19) that mediates their binding to membranes. We examine the roles of various GTPase switch proteins in regulating intracellular signaling and other processes in several later chapters.
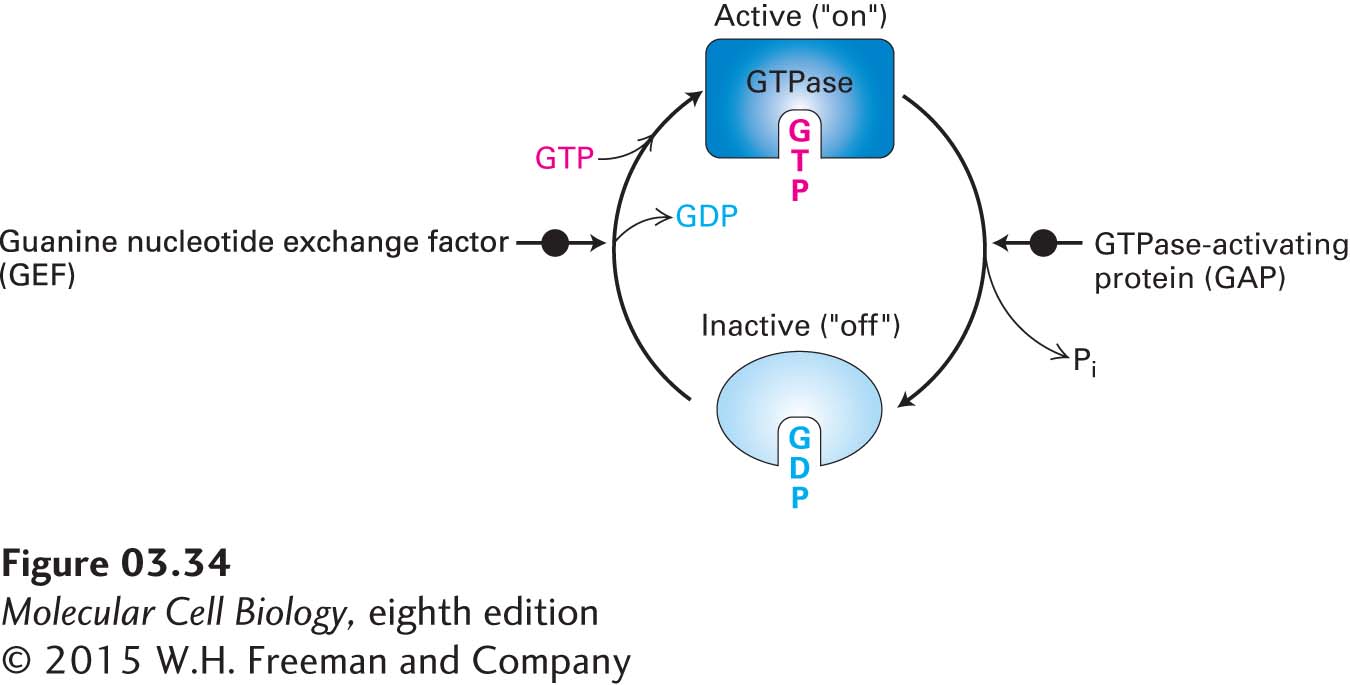
All the GTPase switch proteins exist in two forms, or conformations (Figure 3-34): (1) an active (“on”) form with bound GTP, which can influence the activity of specific target proteins to which they bind, and (2) an inactive (“off”) form with bound GDP. The switch is turned on—that is, the conformation of the protein changes from inactive to active—when a GTP molecule replaces a bound GDP in the inactive conformation. The switch is turned off when the relatively slow GTPase activity of the protein hydrolyzes bound GTP, converting it to GDP and leading the conformation to change to the inactive form. The amount of time any given GTPase switch remains in the active, GTP-bound form depends on the rate of its GTPase activity. Thus the GTPase activity acts as a timer to control this switch. Cells contain a variety of proteins that can modulate the baseline (or intrinsic) rate of GTPase activity for any given GTPase switch and so can control how long the switch remains on. For example, GTPase-activating proteins, or GAPs, increase the rate of GTPase activity, thus reducing the time the GTPase is in the active form. Cells also have specific proteins whose function is to regulate the conversion of inactive GTPases to active ones—that is, to turn the switch on—by mediating the replacement of bound GDP with GTP (GDP/GTP exchange). These proteins are called guanine nucleotide exchange factors, or GEFs. GTPases with lipid anchors are also regulated by proteins called guanine nucleotide dissociation inhibitors (GDIs) that bind to the lipid chain and thus influence interactions with cellular membranes.
FIGURE 3-34The GTPase switch. GTPases are enzymes that bind to GTP and hydrolyze it to GDP. When bound to GTP, the GTPase protein adopts its active, or “on,” conformation and can interact with target proteins to regulate their activities. When the bound GTP is hydrolyzed to GDP by the intrinsic GTPase activity of the protein, the GTPase with GDP bound assumes an inactive, or “off,” conformation. The GTPase switch can be turned back on when another protein, called a GEF (guanine nucleotide exchange factor), mediates the replacement (exchange) of the bound GDP with a GTP molecule from the surrounding fluid. GTPase-activating proteins, or GAPs, can influence the rates of GTP hydrolysis. The binding of the active form of the GTPase to its targets is a form of noncovalent regulation.
The GAPs, GEFs, and GDIs are themselves subject to regulation and, together with their GTPases, participate in complex regulatory networks that control a vast array of cellular activities. It is, therefore, not surprising that disruptions of these finely tuned regulatory networks by mutations or pathogens are associated with a wide variety of diseases. Examples of genetic diseases affecting these networks include Noonan syndrome (a developmental disorder), retinitis pigmentosa (a degenerative eye disease), and X-linked mental retardation. Examples of disruptions of these networks by pathogens include bacterially induced food poisoning, dysenteries (inflammation of the intestines with diarrhea), Legionnaires’ disease (a severe type of pneumonia that involves lung inflammation), and even the plague [also called the Black Death, which between 1347 and 1351 decimated the populations of China (~50 percent death rate) and Europe (~33 percent death rate)].