Although histones are the predominant proteins in chromatin, other, less abundant, nonhistone chromatin-associated proteins, as well as the DNA molecule itself, are also crucial to chromosome structure. Recent results indicate that it is not protein alone that gives a metaphase chromosome its structure. Micromechanical studies of large metaphase chromosomes from newts in the presence of proteases or nucleases indicate that DNA, not protein, is responsible for the mechanical integrity of a metaphase chromosome when it is pulled from its ends. These results are inconsistent with a continuous protein scaffold at the chromosome axis. Rather, the integrity of chromosome structure requires the complete chromatin complex of DNA, histone octamers, and nonhistone chromatin-associated proteins.
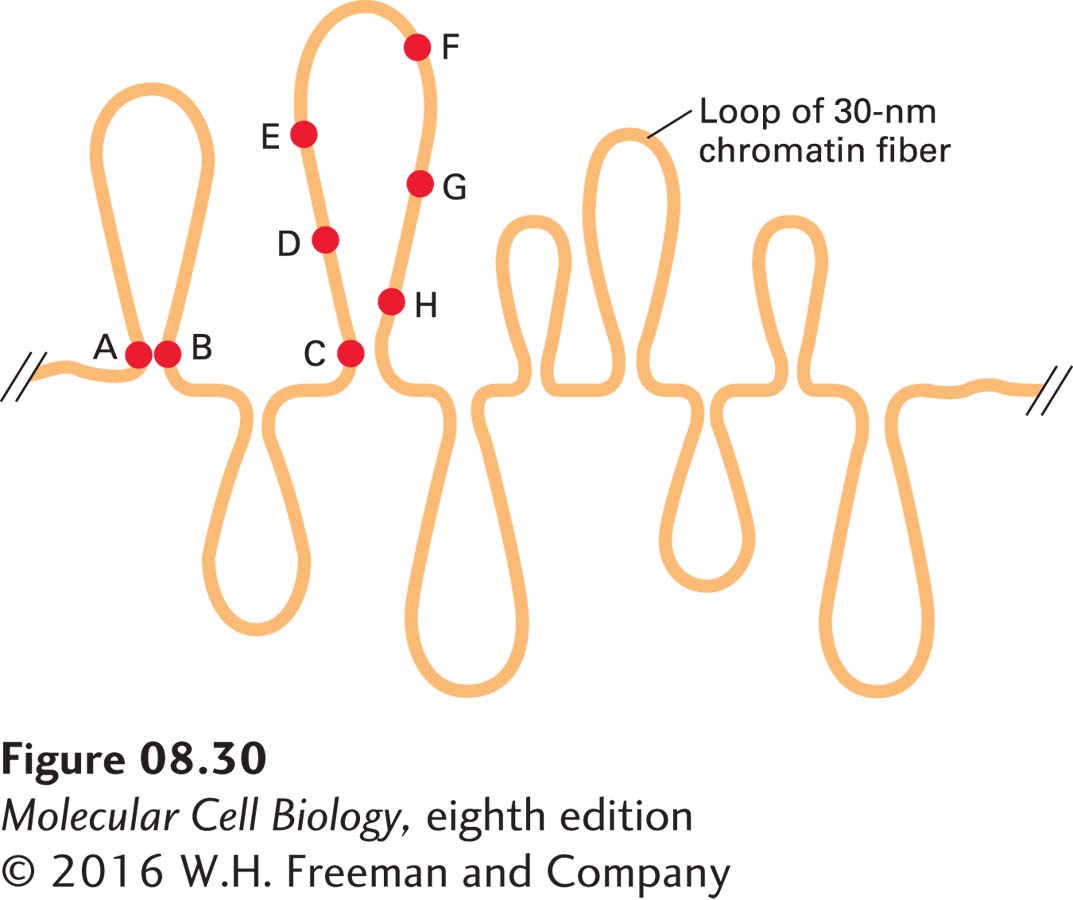
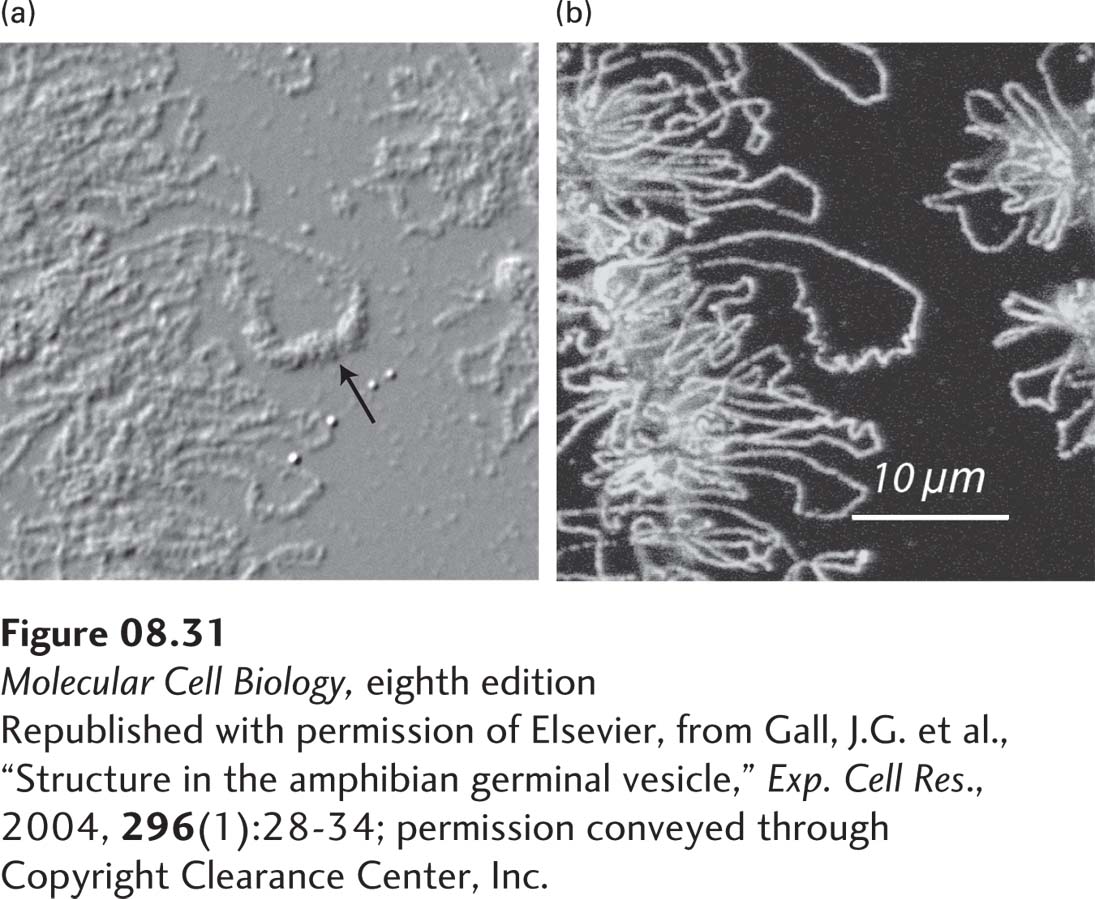
Experiments in which several different fluorescent-labeled probes were hybridized to the DNA of one chromosome in human interphase cells support a model in which chromatin is arranged in large loops. In these experiments, some sequences separated by millions of base pairs in linear DNA appeared very close to one another in interphase nuclei (Figure 8-30). These closely spaced probe sites are postulated to lie at the bases of the chromatin loops. Loops of chromatin can also be directly visualized by light microscopy in the active chromatin of growing amphibian oocytes (“lampbrush chromosomes”) (Figure 8-31). These cells are enormous compared to most cells (~1 mm in diameter) because they stockpile all of the nuclear and cytoplasmic material required for division of the fertilized egg into the thousands of differentiated cells required to generate a feeding embryo that can ingest additional nutrients. The nuclei of these amphibian oocytes are 50 times the diameter of most vertebrate somatic cells. The chromatin of these cells is not constrained by a closely associated nuclear envelope, but rather “floats” in a volume of nucleoplasm about 105 times greater than in most somatic cells. Microscopy of chromatin from these cells stained with antibody to RNA polymerase II shows long loops of chromatin densely packed with polymerase extending from a central core of highly condensed, transcriptionally inactive chromatin. As discussed below, the loops are tethered at their bases by a mechanism that does not interrupt the duplex DNA molecule that extends the entire length of the chromosome.
EXPERIMENTAL FIGURE 8-30Fluorescent-labeled probes hybridized to interphase chromosomes demonstrate chromatin loops and permit their measurement. In situ hybridization to interphase cells was carried out with several different probes specific for sequences separated by known distances in linear, cloned DNA. Lettered circles represent probes. Measurement of the distances between different hybridized probes, which could be distinguished by their color, showed that some sequences (e.g., A and B), separated from one another by millions of base pairs, appear located near one another within nuclei. For some sets of sequences, the measured distances in nuclei between one probe (e.g., C) and sequences successively farther away initially appear to increase (e.g., D, E, and F) and then appear to decrease (e.g., G and H). See H. Yokota et al., 1995, J. Cell Biol.130:1239.
[Republished with permission of Elsevier, from Gall, J.G. et al., “Structure in the amphibian germinal vesicle,” Exp. Cell Res., 2004, 296(1):28-34; permission conveyed through Copyright Clearance Center, Inc.]
EXPERIMENTAL FIGURE 8-31Loop of chromatin in a lampbrush chromosome. A short segment of a lampbrush chromosome in the nucleus of an oocyte from the newt Notophthalmus viridescens. (a) Differential interference contrast (DIC) microscopy of a portion of the lampbrush chromosome and a loop with transcribed RNA associated with hnRNP proteins (arrow; see Chapter 10). (b) The same field observed by immunofluorescence after staining with antibody to RNA polymerase II.
[Republished with permission of Elsevier, from Gall, J.G. et al., “Structure in the amphibian germinal vesicle,” Exp. Cell Res., 2004, 296(1):28-34; permission conveyed through Copyright Clearance Center, Inc.]
Page 336
Ringlike Structure of SMC Protein Complexes The bases of chromatin loops (see Figure 8-30) in interphase chromosomes may be held in place by proteins called structural maintenance of chromosome (SMC) proteins. These nonhistone proteins are critical for maintaining the structure of condensed chromosomes during mitosis. In extracts prepared from the large nuclei of Xenopus laevis (African frog) eggs, chromosomes can be induced to condense as they do in intact cells as they enter prophase of mitosis. This condensation fails to occur when one type of SMC protein is depleted from the extract with specific antibodies. Yeast with mutations in another type of SMC protein fail to properly associate sister chromatids following DNA replication in the S phase. As a result, chromosomes do not properly segregate to daughter cells during mitosis. Related SMC proteins are required for proper segregation of chromosomes in bacteria and archaea, indicating that SMCs are an ancient class of proteins vital to chromosome structure and segregation in all kingdoms of life.
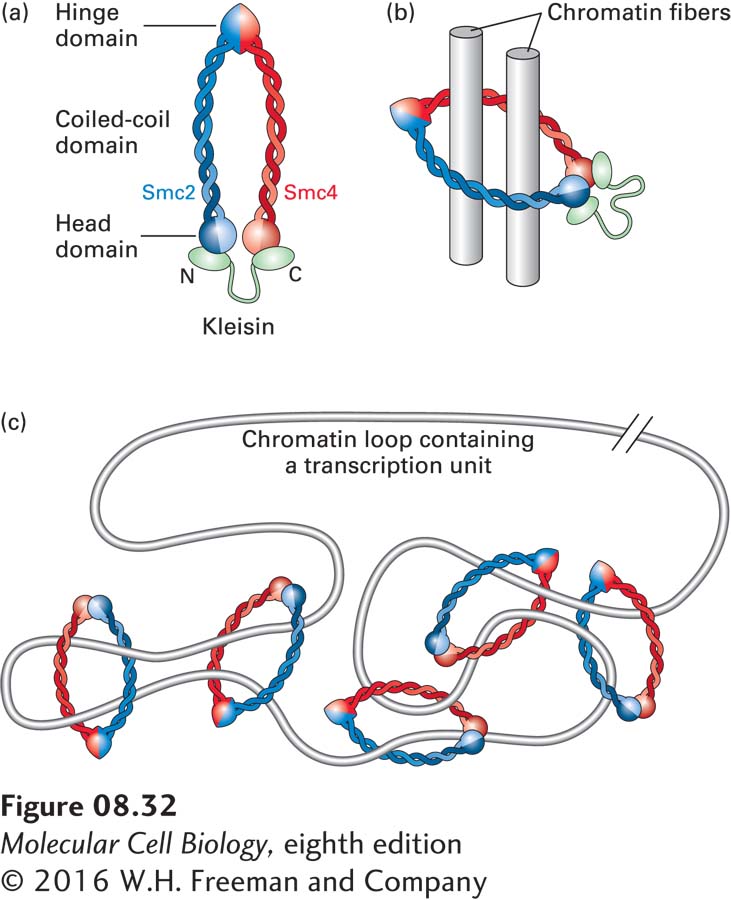
Each SMC monomer contains a hinge region where the polypeptide folds back on itself, forming a very long coiled-coil region and bringing the N- and C-termini together so they can interact to form a globular head domain (Figure 8-32a). The hinge domain of one monomer (blue in Figure 8-32) binds to the hinge domain of a second monomer (red), forming a roughly U-shaped dimeric complex. The head domains of the monomers have ATPase activity and are linked by members of another small protein family called kleisins. The overall SMC complex is a ring with a diameter large enough to accommodate two 30-nm chromatin fibers (Figure 8-32b) and is capable of linking two circular DNA molecules in vitro. SMC proteins are proposed to form the bases of chromatin loops by forming topologically constrained knots in 30-nm chromatin fibers, as diagrammed in Figure 8-32c. This model can explain why cleavage of the DNA at a relatively small number of sites leads to rapid dissolution of condensed metaphase chromosome structure, whereas protease cleavage of proteins has only a minor effect on chromosome structure until most of the protein is digested. When the DNA is cut anywhere in a long region of chromatin containing several chromatin loops, the broken ends can slip through the SMC protein rings, “untying” the topological knots that constrain the loops of chromatin. In contrast, most of the individual rings of SMC proteins must be broken before the topological constraints holding the bases of the loops together are released.
FIGURE 8-32Model of SMC complexes bound to chromatin. (a) Model of an SMC protein complex. (b) Model of SMC complex topologically linking two 30-nm chromatin fibers (represented by cylinders). (c) Model for the binding of SMC complexes to the base of a loop of transcribed chromatin. See K. Nasmyth and C. H. Haering, 2005, Ann. Rev. Biochem.74:595.
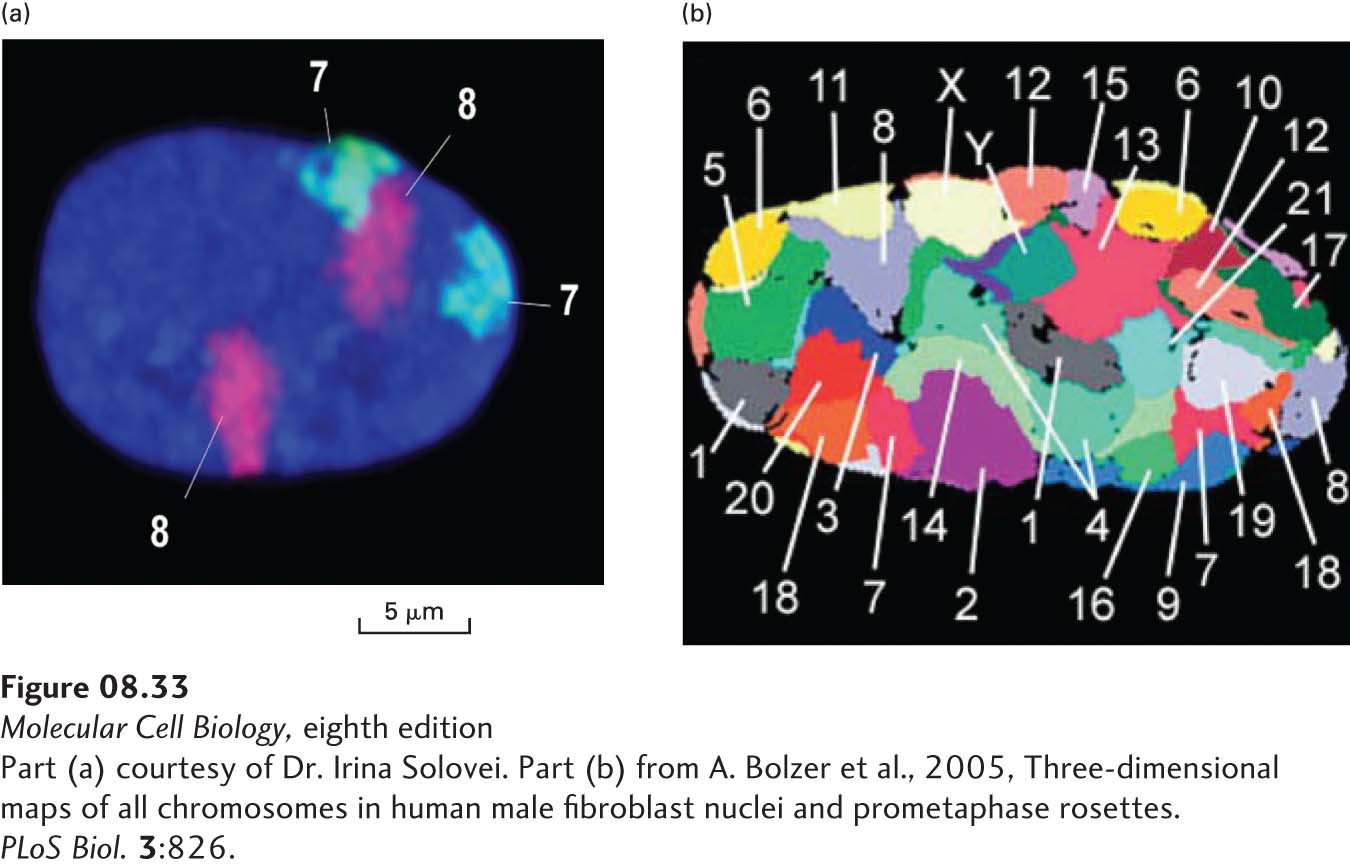
Interphase Chromosome Territories In the small nuclei of most cells, individual interphase chromosomes, which are less condensed than metaphase chromosomes, cannot be resolved by standard microscopy or electron microscopy. Nonetheless, the chromatin of an interphase chromosome is not spread throughout the nucleus. Rather, interphase chromatin is organized into chromosome territories. As illustrated in Figure 8-33, in situ hybridization of interphase nuclei with chromosome-specific fluorescent-labeled probes shows that the probes are localized within restricted regions of the nucleus, rather than appearing throughout the nucleus. Use of probes specific for different chromosomes shows that there is little overlap between chromosomes in interphase nuclei. The precise positions of chromosomes are not reproducible between cells, although large chromosomes tend to lie at the periphery of the nucleus and small chromosomes toward the center. Also, repeats of rRNA transcription units on chromosomes 13, 14, 15, 21, and 22, known as nucleolar organizers, associate with nucleoli found near the center of the nucleus.
[Part (a) courtesy of Dr. Irina Solovei. Part (b) from A. Bolzer et al., 2005, Three-dimensional maps of all chromosomes in human male fibroblast nuclei and prometaphase rosettes. PLoS Biol.3:826.]
EXPERIMENTAL FIGURE 8-33During interphase, human chromosomes remain in non-overlapping territories in the nucleus. (a) Fixed interphase human fibroblasts were hybridized in situ to fluorescently labeled probes specific for sequences along the full length of human chromosomes 7 (cyan) and 8 (purple). DNA is stained blue with DAPI. In the diploid cell, each of the two chromosome 7s and two chromosome 8s is restricted to a territory or domain within the nucleus, rather than stretching throughout the entire nucleus. (b) This image from a fixed interphase fibroblast from a human male was made with a method similar to that used in (a), except that chromosome paint probes specific for each chromosome were hybridized to the cell to reveal the locations of nearly all the chromosomes. Some of the chromosomes are not observed in this confocal slice through the nucleus.
[Part (a) courtesy of Dr. Irina Solovei. Part (b) from A. Bolzer et al., 2005, Three-dimensional maps of all chromosomes in human male fibroblast nuclei and prometaphase rosettes. PLoS Biol.3:826.]
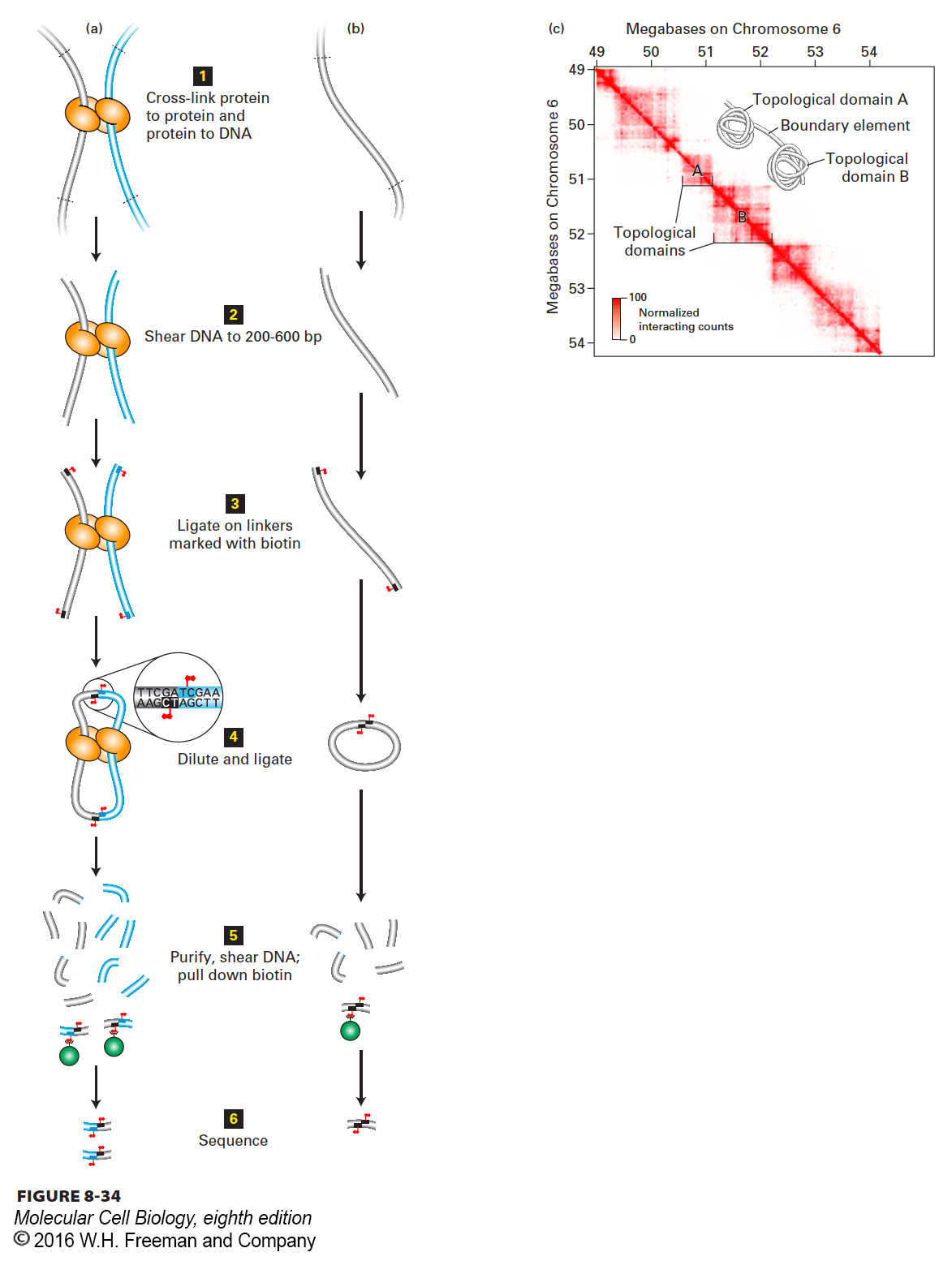
Topological Domains Within Chromosome Territories A group of related methods referred to as chromosome conformation capture (“3C”) methods have been made possible by the advent of massively parallel DNA sequencing and the ability to sequence tens of millions of 50–100-bp DNA fragments (see Chapter 6). The goal of these methods is to determine the three-dimensional spatial organization of chromatin within nuclei of interphase cells. In human interphase cells, about 2 m of DNA is folded and refolded to fit within a nucleus only about 20 µm in diameter! The general strategy of chromosome conformation capture is illustrated in Figure 8-34, where associations between distant regions of chromatin are represented in (a), and regions of chromatin not specifically associated with another region are shown in (b). First (step 1), intact cells are treated with chemical cross-linkers such as formaldehyde that diffuse through cell membranes and covalently cross-link protein to protein and protein to DNA. Next 2, the cross-linked chromatin is isolated and either digested with a restriction enzyme or subjected to intense sonication to mechanically shear the DNA into fragments of 200–600 bp. 3 Short oligonucleotide linkers are then ligated to the ends of the DNA fragments. These linkers contain a cytosine linked to a biotin at the 5′ position (see Figure 2-17). 4 After removal of excess linker, the preparation is diluted considerably and subjected to treatment with DNA ligase. Because the preparation is diluted, ligations occur preferentially between the ends of fragments held in close proximity by cross-links. 5 After ligation, the protein-DNA cross-linking reaction is reversed, the protein is digested with proteases, and the DNA is isolated and further sonicated. Ligated fragments are separated from other fragments using streptavidin, which binds the biotin in the linkers added in step 3.
[Part (c) data from J. R. Dixon, 2012, Nature485:376.]
FIGURE 8-34Chromosome conformation capture. (a, b) Strategy of chromatin conformation capture methods. See text for explanation. Gray and blue tubes represent regions of DNA separated by >10 kb in the genome sequence. Biotin is represented by red flags. See E. Lieberman-Aiden, 2009, Science326:289. (c) Heat map of chromosome conformation capture data for a region of chromosome 6 in mouse embryonic stem cells. The sequence from 49 to 54 Mb from the left end of chromosome 6 is represented on both axes. Each pixel shows data from a 10 kb sequence. The number of times a sequence from one 10-kb region indicated on the x axis was ligated to a sequence from a second 10-kb region on the y axis is indicated by the intensity of red color, as shown in the key at the lower left. A value of 100 (dark red) indicates that a sequence anywhere within the 10-kb region on the x axis was found ligated to a sequence from anywhere in the 10-kb region on the y axis 100 times. Since the probability that two ends generated by sonication will be ligated together is higher for ends that are close together than for ends that are far apart, the intensity of the red color in any pixel indicates the relative proximity of the sequences in the two 10-kb intervals in the nuclei at the time of cross-linking. Inset shows a model of chromatin folding that is consistent with these results.
[Part (c) data from J. R. Dixon, 2012, Nature485:376.]
Page 337
The purified, ligated fragments are then sequenced. Ligation points are marked by the duplicated sequence of the oligonucleotide linker (see Figure 8-34, step 4). Sequences on each side of the ligation point are mapped to the genome. In cases in which the two ends of a single fragment were linked together (Figure 8-34b), the sequences on either side of the oligonucleotide linker sequence will map within a few hundred bases of each other on the genome, since the ligated fragments were only a few hundred bases long. But in cases in which fragments that are distant in the genomic sequence were ligated together because they were cross-linked to proteins holding them together (Figure 8-34a), the sequences will map far apart. The observation of distant (>10 kb) sequences repeatedly ligated to each other implies that their respective regions of chromatin were associated with each other in vivo.
The data from a chromatin conformation capture assay can be plotted on a two-dimensional heat map (Figure 8-34c). In this plot, the sequence of the same portion of the genome is plotted on both the x and y axes, with each pixel equivalent to 10 kb. The pixel at coordinates x, y is colored red where a sequence at coordinate x was ligated to a sequence at coordinate y. The intensity of the red color is proportional to the numbers of ligation events observed that linked a sequence in the 10-kb interval x with a sequence in the 10-kb interval y.
Page 338
Figure 8-34c shows the plot generated for a roughly 5.5-Mb region of chromosome 6 in mouse embryonic stem cells. It is immediately apparent that the genome is divided into regions called topological domains, in which a chromosomal region is far more likely to be ligated to another sequence within the same topological domain than it is to be ligated to a sequence in another topological domain. These topological domains are on the order of 200 kb to 1.5 Mb in length, with a median size of 880 kb. For example, sequences in the interval of chromosome 6 between 50.9 Mb and 51.3 Mb (see Figure 8-34c, topological domain A) are much more likely to be ligated to each other than to sequences in the interval from 51.3 Mb to 52.2 Mb (topological domain B), or to sequences from any of the other topological domains that are apparent. In situ hybridization studies showed that sequences within a topological domain lie much closer to each other in the fixed cell nucleus than to sequences the same distance away in base pairs, but in a neighboring topological domain. These results have been interpreted to indicate that the chromatin fiber is folded into topological domains, as represented in the inset of Figure 8-34c. The topological domains are separated by shorter regions of chromatin, called boundary elements, that do not interact with distant regions of chromatin. Since the topological domains are on the order of 200 kb–1.5 Mb in length, they are long enough to contain several average-sized genes. The topological domains identified by these chromatin conformation capture assays may correspond to the loops of chromatin observed in the lampbrush chromosomes described above, which are not constrained by the nuclear envelope of a vastly smaller nucleus and have an opportunity to unfold (see Figure 8-31). Current research is exploring what protein-DNA interactions might be responsible for establishing boundary elements between topological domains. As we will see in Chapter 9, related chromosome conformation capture techniques have provided strong evidence that proteins bound to enhancers interact with proteins bound to promoters many kilobases away.
Page 339
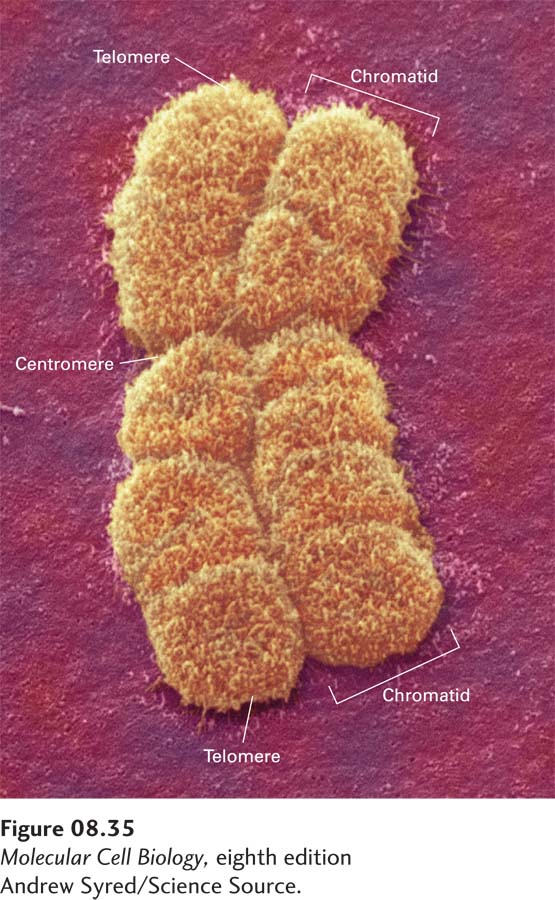
Metaphase Chromosome Structure Condensation of chromosomes during prophase (see Figure 18-37) may involve the formation of many more loops of chromatin, so that the length of each loop is greatly reduced compared with chromatin loops in interphase cells. As a result, chromosomes condense into structures of much greater width than interphase chromosomes and decrease in length severalfold, generating the condensed chromosomes observed during metaphase (Figure 8-35).
[Andrew Syred/Science Source.]
FIGURE 8-35Typical metaphase chromosome. As seen in this scanning electron micrograph, the chromosome has replicated and comprises two chromatids, each containing one of two identical DNA molecules. The centromere, where the chromatids are attached at a constriction, is required for their separation late in mitosis. Special telomere sequences at the ends function in preventing chromosome shortening.
[Andrew Syred/Science Source.]
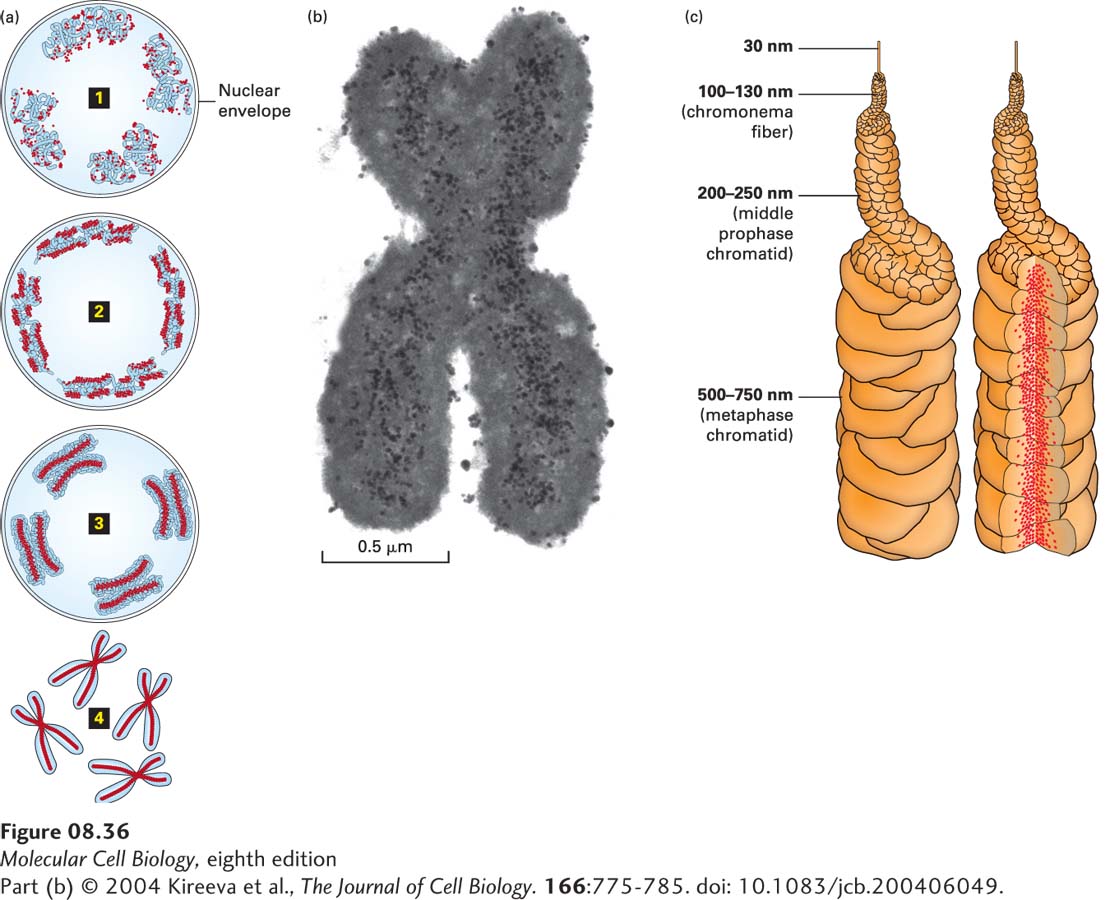
The geometry of chromatin in metaphase chromosomes is not well understood. Experiments with frog egg extracts have shown that a protein complex called condensin, composed of SMC subunits (see Figure 8-32 and Chapter 19), contributes to chromosome condensation using energy from ATP hydrolysis. Microscopic analysis of mammalian chromosomes as they condense during prophase indicates that in the initial period of prophase, the 30-nm chromatin fiber folds into a 100–130-nm chromonema fiber associated with the nuclear envelope (Figure 8-36). Chromonema fibers then fold into structures with a diameter of 200–250 nm, called middle prophase chromatids (Figure 8-36a, 3), which then fold into the 500–750-nm-diameter chromatids observed during metaphase when the nuclear envelope retracts into the endoplasmic reticulum (Figure 8-36a, 4) (see also Chapter 19). Ultimately, the full lengths of the two associated daughter chromosomes generated by DNA replication during the previous S phase of the cell cycle (see Figure 1-21) condense into bar-shaped structures (chromatids) that in most eukaryotes are linked at the central constriction called the centromere (see Figure 8-35). An electron micrograph of a section through a metaphase chromosome stained with anti-SMC antibodies linked to small gold spheres (Figure 8-36b) shows that condensin, proposed to be at the bases of chromatin loops (see Figure 8-32c), occupies approximately one-third of the chromatid diameter (Figure 8-36c, right), where it contributes to the shaping of each chromatid.
FIGURE 8-36Model for mitotic chromosome condensation. (a) Stages of chromosome condensation during mitosis. Changes in large-scale chromatin folding (blue) versus distribution of Smc2, a subunit of condensin (red), from early prophase 1 to middle prophase 2 to late prophase 3 to metaphase 4. (b) Transmission electron micrograph of immunogold staining of Smc2 in a section through a metaphase chromosome reveals axial staining of Smc2 of about 0.15–0.2 µm in width. (c) “Hierarchical folding, axial glue” model of metaphase chromosome structure. (Left) 30-nm fiber folds into 100–130-nm chromonema fiber, which folds into 200–250-nm middle prophase chromatid, which folds into 500–750-nm metaphase chromatid. Only one chromatid is shown. (Right) Axial condensin distribution (red) occupies approximately one-third of the chromatid diameter, acting as a cross-linking “glue” to stabilize the structure of the metaphase chromosome.