CONCEPT38.1 Fertilization Activates Development
The development of an animal, a plant, or a fungus is one of the most breathtaking processes you will encounter in your study of biology. In sexually reproducing animals, one cell, the fertilized egg, gives rise to an entire organism. Early investigators assumed that a tiny person was encapsulated in a human egg or sperm and that this “homunculus” required only the appropriate stimulus and nutrients for growth:
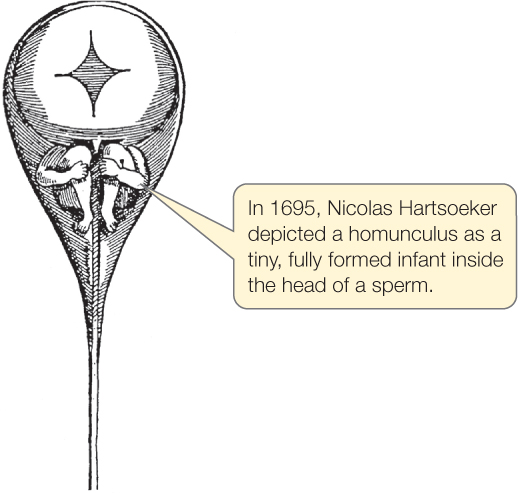
Improvement in microscopes allowed scientists to see that no tiny animal resides in either sperm or egg. That left the mystery—how does an organism achieve its final form?
You saw earlier that scientists obtain much information by studying “model” organisms (see Concept 1.1). Nowhere is this more true than in developmental biology. It is important to realize that certain animals are used as models, not because they represent the majority of the group under investigation, but because of particular characteristics that make them desirable for research, such as short developmental times and easy maintenance in a laboratory. Remember, too, that there are probably millions of animal species—many not yet discovered. Each has tweaked its development to adapt to its environment. The doors are open for further investigations in developmental biology.
Egg and sperm make different contributions to the zygote
Development usually begins with the union of egg and sperm to form a fertilized egg, or zygote. Egg cells are very large compared with other cells—and compared with sperm, they are gigantic. Even the non-yolky eggs of sea urchins and mammals are large compared with other cells, because they contain the materials and information needed to initiate and maintain early development. The cytoplasm of an egg contains most of the molecular machinery for cell division, energy production, and growth (yolk that provides nutrients for growth, mitochondria, ribosomes, etc.), as well as informational molecules such as transcription factors and messenger RNAs (mRNAs). These play important roles in setting up the signaling cascades that orchestrate the major processes of development discussed in Concept 14.1: determination, differentiation, morphogenesis, and growth.
The union of egg and sperm is complex. Many sperm are attracted to an egg. However, one sperm, and only one, must convey its nuclear material to the egg cytoplasm:
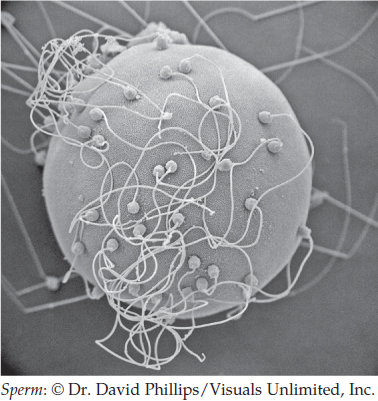
All of the egg cell membrane may be available for sperm entry, as in sea urchins, or entry may be restricted to certain parts. In amphibians, for example, sperm can penetrate only half the egg surface. In Drosophila, sperm must enter through a tunnel whose position is fixed prior to fertilization.
Activation of an egg by a sperm brings about changes in the egg cell membrane and the release of ions from stores within the egg cytoplasm. These ion changes trigger a series of crucial events. Entry of additional sperm is blocked, metabolic rate rises, protein synthesis is initiated, and cytoplasm is reorganized.
Most of the sperm cytoplasm is discarded when the egg and sperm membranes fuse, but the nuclear material and usually the centrosome (the major microtubule organizing center of an animal cell) are transferred from the sperm to the egg. The centrosome then plays a major role in forming the zygote centrosome and organizing the zygote’s microtubules. When egg and sperm nuclei join, the fertilized egg becomes a single-celled, diploid zygote that is ready for its first mitotic division.
LINK
Concept 37.1 explains sperm–egg recognition and blocks to polyspermy (fertilization by more than one sperm)
Polarity is established early in development
Most animals have a defined head and tail, back and belly, and right and left sides. These differences are known as polarities and are usually established early in development. In most animals, polarities are established during early cell divisions as materials deposited in the egg cytoplasm by the mother are redistributed and segregated. In some animals, however, polarities are set in place before or at fertilization.
In the fruit fly Drosophila melanogaster, a favorite model organism, the position of the egg in the ovary determines both anterior–posterior (head–tail) and dorsal–ventral (back–belly) polarity. Cells that surround the egg, called follicle cells, pass mRNAs from the mother to specific regions of the egg cytoplasm. After the egg is fertilized and development is underway, these mRNAs encode regulatory proteins that determine polarity.
789
In amphibians, another group of favorite model organisms, anterior–posterior and dorsal–ventral polarity are determined at fertilization by the position of the yolk and the point of sperm penetration. Cytoplasmic rearrangement following fertilization is easily observed in some frog species. In unfertilized eggs of these species, dense yolk granules are concentrated by gravity in the lower half of each egg, the vegetal hemisphere. This yolky vegetal hemisphere is unpigmented. In the top half of each egg, or animal hemisphere, the outermost (cortical) cytoplasm is heavily pigmented, while the pigment in the underlying cytoplasm is more diffuse. Sperm-binding sites occur only on the animal hemisphere. When a sperm enters the egg, the cortical cytoplasm rotates toward the site of sperm entry, producing a band of lightly pigmented cytoplasm called the gray crescent on the opposite side (FIGURE 38.1).
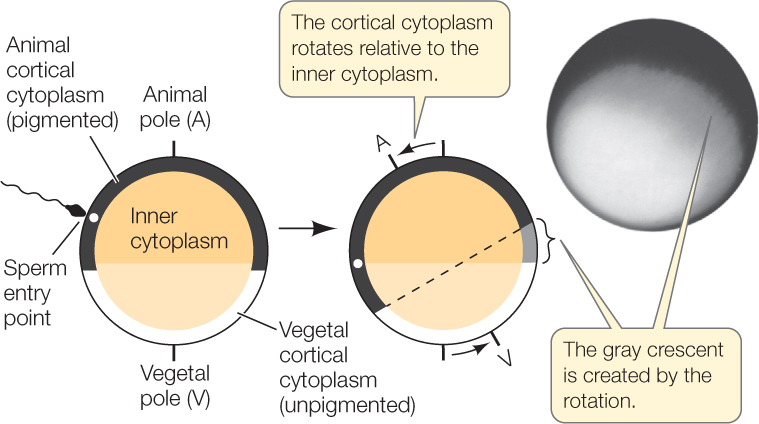
As we will soon see, the anus later forms within the gray crescent. Thus with the formation of the gray crescent, the zygote already has an anterior–posterior axis for a future head and tail, as well as a dorsal–ventral axis. The cytoplasmic rearrangement is initiated by the sperm centriole, which organizes microtubules in the egg cytoplasm in a parallel array that extends through the vegetal hemisphere. This cytoskeleton guides the shift of the cortical cytoplasm (see Concept 14.2), and the gray crescent appears.
Much of our understanding of developmental mechanisms builds on the insightful experiments of scientists in the late 1800s and early 1900s who used techniques that may seem primitive by today’s standards but were very revealing. This point is beautifully illustrated by German biologist Hans Spemann’s work, which also had a profound influence on future amphibian research. Similar experiments were carried out on many animals, and today’s work is based on the results of these creative early investigators.
In the early 1900s, Spemann was studying the development of salamander eggs. With great patience and dexterity, he formed loops from single hairs taken from his baby daughter (baby hair is finer than adult hair) and tied them around fertilized eggs, effectively dividing the eggs in half (FIGURE 38.2). When his loops bisected the gray crescent, both halves of the zygote developed into a complete embryo. When he tied the loops so the gray crescent was on only one side of the constriction, only the half with the gray crescent developed normally. The half lacking gray crescent material became an unorganized clump of cells, which, because of its characteristics, he referred to as the “belly piece.” Spemann concluded that important cytoplasmic factors—now called cytoplasmic determinants—necessary for the establishment of polarity and development of a normal salamander are unequally distributed in the fertilized egg and reside in the gray crescent.
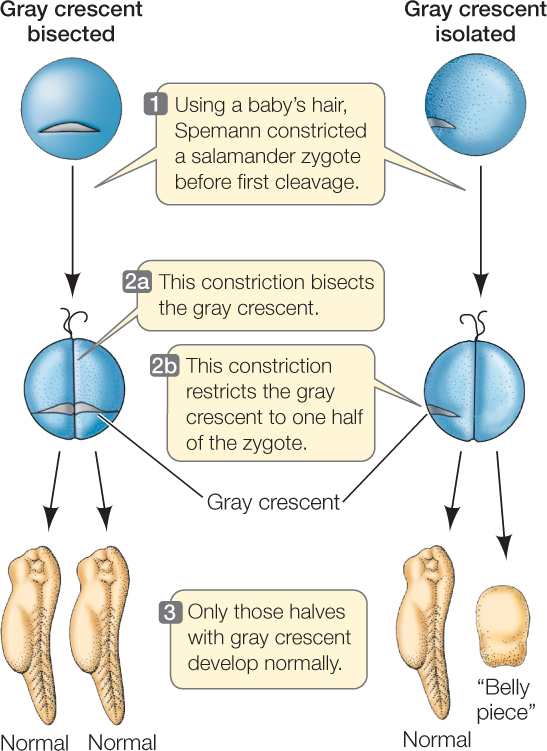
Experiments using amphibian eggs have contributed much to our understanding of development. Keep in mind, though, that although amphibians are useful models, many animals develop differently, and in many, polarities are not established until cell divisions are underway.
CHECKpointCONCEPT38.1
- What normally initiates development?
- What does the sperm contribute to the zygote?
- What usually determines initial polarity in developing embryos?
- How are anterior–posterior and dorsal–ventral polarities established in fruit flies? In frogs?
790
We have now seen how the zygote forms and how polarities may be established. We will turn next to the early series of cell divisions that produce a multicellular embryo.