1.1 THE EVOLUTION OF LIFE ON EARTH
All organisms on Earth are connected by an evolutionary journey spanning more than 3 billion years. The diversity of life we see around us is the sum of a limitless number of mutations, changes in genetic information that are usually subtle but sometimes dramatic. When Charles Darwin proposed that natural selection acts on variation in populations, he had no knowledge of the mechanisms that give rise to that variation. Such mechanisms lie at the heart of modern molecular biology.
What Is Life?
Almost anyone can distinguish a living organism from an inanimate object. However, a rigorous scientific description of life is harder to achieve. Life differs from nonlife in identifiable ways, as summarized in Figure 1-1. Organisms move, reproduce, grow, and alter their environment in ways that inanimate objects cannot. But such characteristics alone provide an unsatisfying definition of life, particularly when a few of them may be shared by inanimate substances. In 1994, the United States National Aeronautics and Space Administration (NASA) convened a panel to consider the question, “What is life?” A simple definition resulted: Life is a chemical system capable of Darwinian evolution. The importance of evolutionary theory to all biological sciences gains full expression in this concise statement.
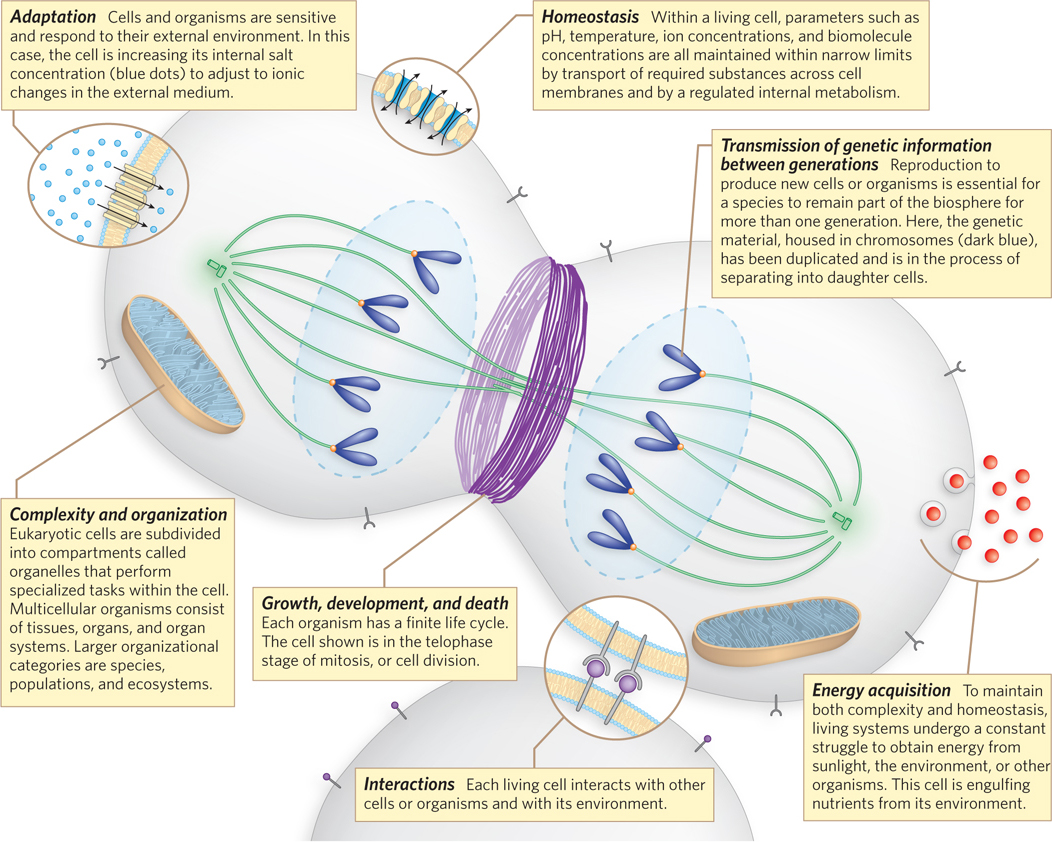
Every living system we know about has several requirements for its existence. Two of these—
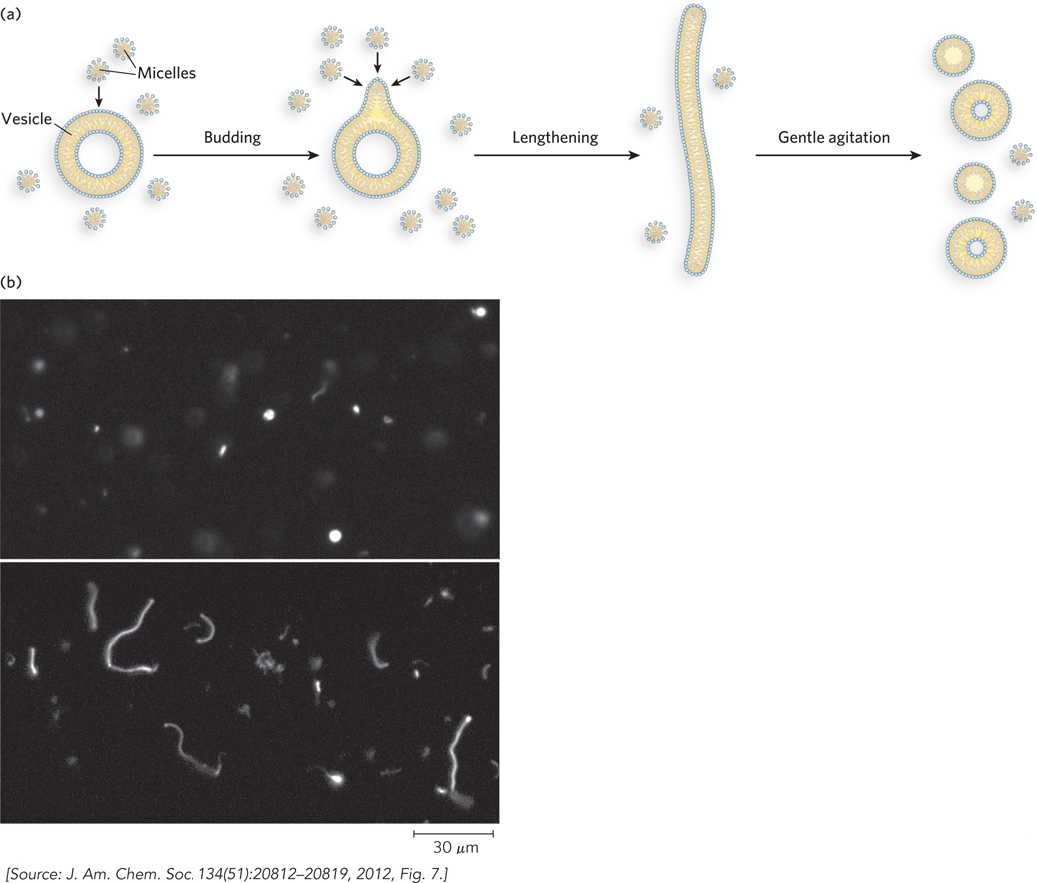
A third requirement for a living system is an envelope, creating a barrier between the living and inanimate worlds and establishing a means of selective interaction between a cell and its environment. The work of Jack Szostak, chronicled in this chapter’s Moment of Discovery, may be replicating some key evolutionary moments that led to enveloped living systems (Figure 1-2).
3
The final two requirements—
The presence of enzymes in a cell depends on the faithful transmission of the genetic information that encodes them from one generation to the next. Enzymes, as well as the myriad other proteins and RNA molecules that regulate their synthesis and function, are the actual molecular targets of evolution. When a cell acquires a new function, it generally reflects the presence of a new enzyme or set of enzymes, or an alteration in the regulation or function of an existing enzyme or process. The new functions arise through changes in genes—
4
Evolution Underpins Molecular Biology
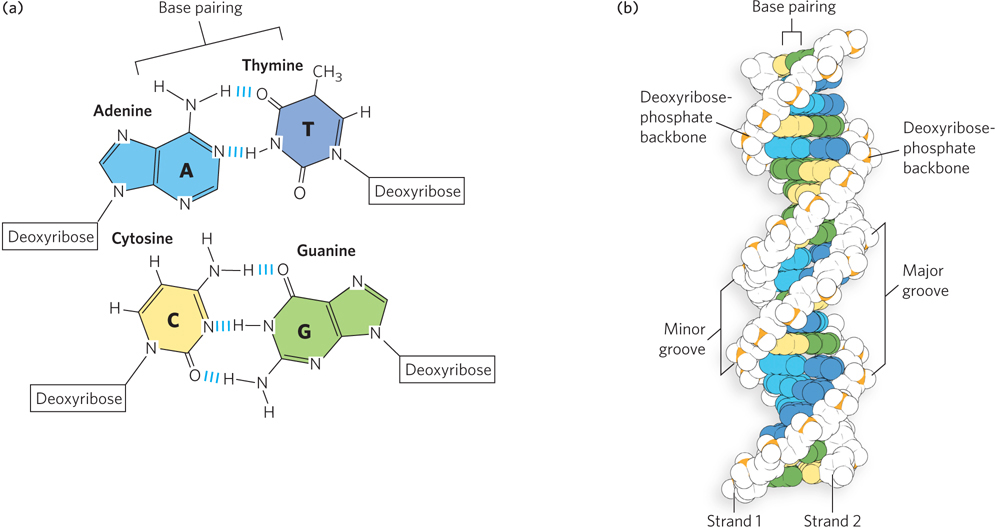
In 1973, the geneticist Theodosius Dobzhansky published an article in the professional journal The American Biology Teacher entitled “Nothing in Biology Makes Sense Except in the Light of Evolution.” This sentiment has special meaning in molecular biology, because the pathways and processes in living systems give rise to the genetic variation on which natural selection acts (Figure 1-4). They also inform the ongoing investigations into how life arose on Earth.
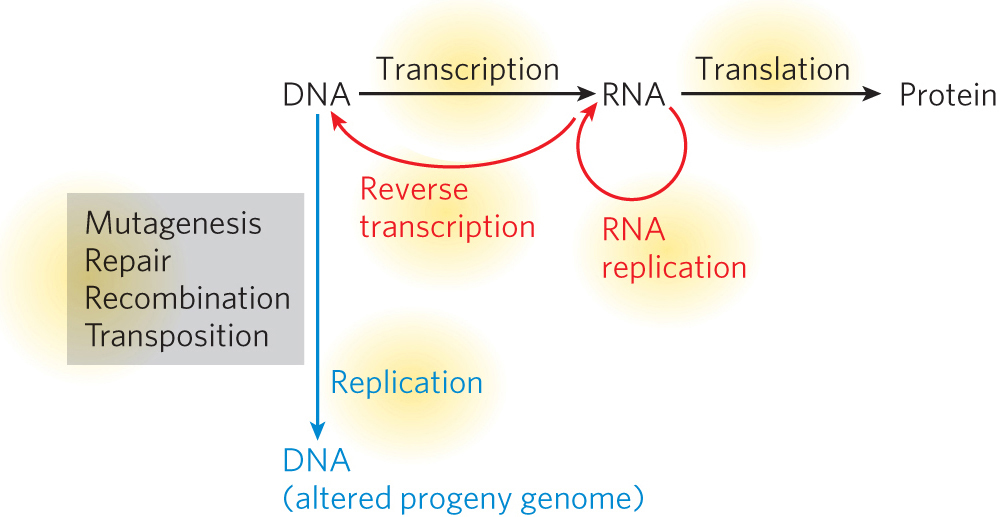
Evolution relies on spontaneous and generally random changes in an organism’s genomic material, called mutations. In spite of the elaborate cellular mechanisms we consider in this book, all of which help ensure accurate transmission of genetic information from one generation to the next, mutations regularly occur. Mutations can be as simple as a change in a single base pair of DNA or base of RNA or as substantial as the inversion, deletion, or insertion of large segments of genetic material. As we will be discussing in detail, errors can arise during replication (Chapter 11), and DNA damage can lead to permanent mutation when repair systems (Chapter 12) go awry. Larger chromosomal changes can arise from recombination (Chapter 13) or transposition (Chapter 14). Some mutations affect genes directly; others affect the ways in which DNA is transcribed into RNA or RNA is processed or translated (Chapters 15, 16, 17 and 18). Relatively minor changes in genes involved in regulatory processes (Chapters 19, 20, 21 and 22) can give rise to dramatic changes in the organism; this realization has created a new field, essentially a modern merger of the fields of evolutionary and developmental biology, dubbed “evo-
5
An understanding of these processes has also given us insights into the origins of life and the process of evolution. Continuing explorations of RNA structure (Chapter 6) and metabolism (Chapters 15 and 16) have informed new theories of prebiotic evolution. The genetic code (Chapter 17) provides a particularly vivid look at the shared history of every organism on Earth.
Molecular biology has provided the enzymes that make most of the methods of biotechnology possible (Chapter 7). These increasingly powerful methods for studying the genes of many different organisms allow us to trace their evolution. Through modern genomics (Chapter 8), molecular biology is opening a window onto evolution that Charles Darwin would marvel at.
The interrelationship of molecular biology and evolution is of more than academic interest. Human beings exist in a world where every organism continues to evolve. Microorganisms, with their short life cycles, evolve most rapidly (Highlight 1-1). Of special concern are human pathogens, as well as the microorganisms, fungi, insects, and other organisms that affect our food crops, livestock, and water supply. Molecular biology provides essential tools for use in tracking pandemics, investigating new microbial pathogens, identifying the genes underlying human genetic diseases, solving crimes, tracing the origin of diseases, treating cancer, and engineering microorganisms for new purposes in bioremediation and bioenergy. All of these efforts rely heavily on the concepts of evolutionary biology. Indeed, modern society relies on countless innovations in medicine and agriculture that would not exist but for Darwin’s great insight.
6
HIGHLIGHT 1-1 EVOLUTION: Observing Evolution in the Laboratory
The bacterium Deinococcus radiodurans has a remarkable capacity to survive the effects of ionizing radiation (IR, or gamma rays). A human being would be killed by exposure to 2 Gy (1 Gy (gray) = 100 rads) of IR, but cultures of Deinococcus routinely survive 5,000 Gy with no lethality. Deinococcus is a desert dweller, and this characteristic reflects its adaptation to the effects of desiccation. In dry conditions, the bacterium cannot grow and its cellular metabolism shuts down. Spontaneous damage to the cellular DNA accumulates, including strand breaks. DNA repair processes, which require ATP generated by cellular metabolism, do not take place. However, the bacterium can repair its genome quickly when conditions favorable for growth return. Like desiccation, IR also generates numerous DNA strand breaks, and that same extraordinary capacity for DNA repair is put to use after exposure to IR.
How long does it take for a bacterium to evolve extreme resistance to IR? A recent study demonstrated that Escherichia coli, the common laboratory bacterium, can acquire this resistance by directed evolution. Twenty cycles of exposure to enough IR to kill more than 99% of the cells, with each cycle followed by the outgrowth of survivors, produced an E. coli population with a radiation resistance approaching that of Deinococcus. The entire selection process can be achieved in less than a month. Complete genomic sequencing of cells isolated from the evolved populations typically reveals 40 to 80 mutations. The answer to survival varies from cell to cell, with different cells displaying different arrays of mutations, even when they come from the same evolved population. In just a single, small bacterial culture, evolution can take many paths, and a variety of solutions are found that lead to acquisition of a new trait.
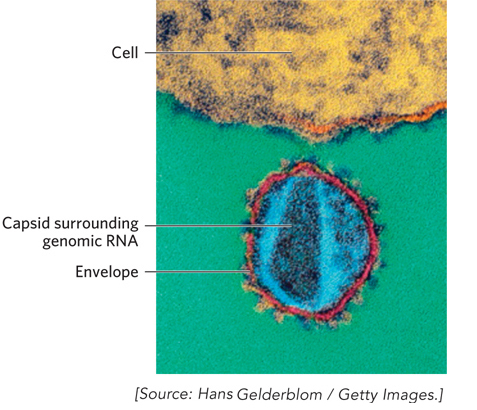
This is just one of many experiments demonstrating that dramatic changes in microorganisms can be readily generated and observed in the laboratory within short periods of time. The same kind of evolutionary processes are occurring constantly in microorganisms in our environment, including human pathogens. When AIDS appeared as a new threat to human health in the early 1980s, the power of evolutionary theory was quickly on display. The causative agent, HIV, was soon isolated and its genomic sequence determined. Characterizing this novel and very dangerous virus from scratch would have delayed treatments for years. But scientists had a shortcut at hand. A deep reservoir of information about viruses and their evolutionary relationships had already been built up over decades of research. The small HIV genome thus held all the clues that science needed for a rapid understanding of its infection cycle and the development of a medical response. Its genome revealed that HIV is a type of RNA virus called a retrovirus, with clear evolutionary relationships to other viruses that were already known and understood (Figure 1). It was immediately evident which HIV genes encode the enzymes essential to the virus life cycle, and these enzymes rapidly became drug targets. One result was the development of highly effective treatments at an unprecedented rate, ranging from AZT to protease inhibitors (see Highlights 5-2 and 14-3 for more detailed descriptions of the retrovirus life cycle). Millions of lives have been saved, in large measure because all biological and medical research is carried out in the context of evolutionary theory.
Life on Earth Probably Began with RNA
About 4.6 billion years ago, the sun and Earth and the other planets and asteroids of our solar system were formed. Within the first billion years of our planet’s existence, life appeared on its surface. How did this happen, and how likely is it that this has happened on other, similar worlds? Modern geologists, paleontologists, and molecular biologists are slowly piecing together the history of life on Earth from the rich trove of clues in the geologic, fossil, and genomic records. A plausible sequence emerges, providing a wide range of hypotheses that can be tested using modern chemical and physical methods.
7
The first few hundred million years were a time of prebiotic chemistry (Figure 1-5). No life was present, but chemical reactions were happening everywhere. The atmosphere contained primarily water, methane, ammonia, hydrogen, nitrogen, and carbon dioxide. Reactions driven by the constant stream of energy coming from the sun were slowly yielding more complex molecules such as simple sugars, amino acids, and nucleotide bases. And the accumulation of organic material was supplemented by materials from a multitude of collisions between early Earth and meteors laden with organic matter. Prebiotic chemistry is being studied by a large community of researchers. A small sampling of their work is presented in the How We Know section at the end of this chapter.
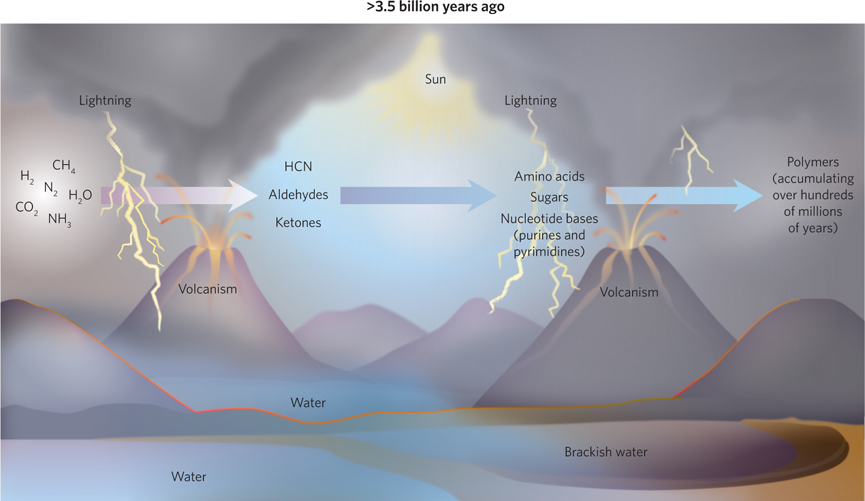
Over a period of millions of years, the accumulation of reaction products yielded a soup containing molecules and polymers. As they grew increasingly complex, particular polymers acquired the capacity to duplicate themselves. The first self-
We do not know what this first “living” polymer was. However, modern molecular biology has given us many reasons to think that RNA either was the first self-
The RNA world hypothesis was first proposed as a stage in evolution by molecular biologists Carl Woese, Francis Crick, and Leslie Orgel, in separate papers published in the late 1960s. The hypothesis describes a living system (or set of living systems) based on RNA. In this system, a variety of RNA enzymes could catalyze all of the reactions needed to synthesize the molecules required for life from simpler molecules available in the environment. The RNA enzymes would include replicators to duplicate all of the RNA catalysts. The “RNA organism,” out of equilibrium with its surroundings, would have to be defined by a boundary. The experiments of Szostak and colleagues show one way in which lipid-
Four more-
8
What is the real catalytic potential of RNA? The second line of supportive research demonstrated that RNA molecules generated in the laboratory can catalyze almost any imaginable reaction needed in a living system—
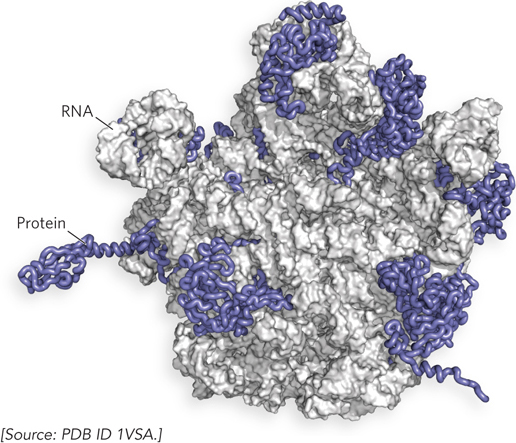
The third and fourth discoveries have further broadened our perspectives on RNA function. We now know that in ribosomes, the large ribonucleoprotein complexes that translate RNA into protein, the RNA is the active component with the capacity to catalyze protein synthesis (Figure 1-6; see also the Moment of Discovery for Chapter 18). Finally, and most recently, RNA sequences capable of simple forms of self-
Ongoing research thus makes it possible to visualize a highly plausible sequence of events unfolding on the pathway from prebiotic soup to living systems. Arising from a myriad random primordial polymers, an RNA world came into being and gradually became more complex. An RNA capable of reliable self-
The Last Universal Common Ancestor Is the Root of the Tree of Life
Countless nascent life forms probably arose from the primordial soup, along with many biological advances that improved their fitness. Successful combinations of RNA catalysts gave way to systems based on protein catalysts. Improvements in catalytic efficiency appeared, along with systematized genetic codes to link genetic information in RNA and DNA to protein sequences. Additional changes facilitated cellular metabolism and reproduction. Protein synthesis was systematized through the evolution of an efficient ribosome machine. RNA became more specialized for information storage and transmission. Cell membranes became more structured and specialized, eventually including mechanisms to selectively transport materials into and out of the cell as needed. And some processes became regulated. In this way, a variety of primitive cells may have evolved—
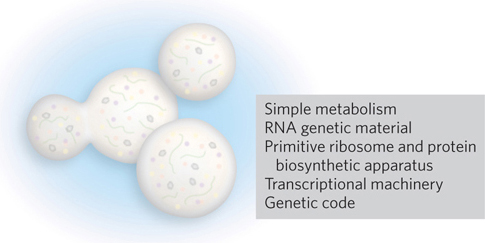
LUCA is a special source of fascination for molecular biologists. Although LUCA probably lived more than 3 billion years ago, our speculation about what this cell was like is informed by experiment. One approach is to determine the minimum protein and genetic requirements for life. Attempts to create a minimal life form, either by reconstituting basic components or by taking bacteria and tsripping them of all unnecessary parts, are underway in laboratories around the world. These experiments are not only defining properties that must have been present in LUCA; they are also setting the stage for the laboratory generation of engineered bacterial cells that can be used to manufacture chemicals for bioenergy, agriculture, and medicine.
9
Another approach to understanding LUCA is to survey all types of living systems on Earth to determine which genes or characteristics are universal. The only genes that are truly universal in living systems are those encoding the cellular machinery for protein synthesis and some components of RNA transcription. All organisms also share (with very minor modifications discussed in Chapter 17) the same genetic code. That same code must have been present in LUCA. To support protein synthesis and RNA synthesis, a simple metabolism must have been present that allowed the uptake of chemical energy and its use to synthesize amino acids, nucleotides, and whatever lipids existed in the cell membrane from precursors available in the environment. The study of LUCA is described in more detail in Chapter 8.
The appearance of LUCA signaled the beginning of biological evolution on Earth. New types of cells gradually appeared, and new environments were exploited. The first cells were capable of taking up organic molecules from their surroundings and converting them to the molecules needed to support protein and RNA synthesis. Cellular complexity resulted in ever-
The early single-
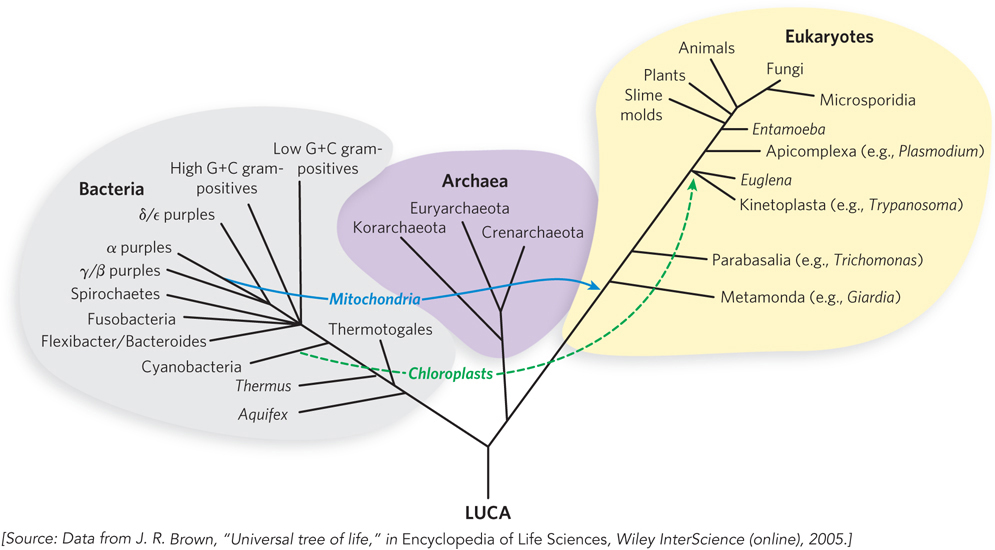
Many additional events helped shape the life we see around us. Notably, photosynthesis appeared about 2.5 billion years ago, as evidenced by the sudden rise in the concentration of atmospheric oxygen documented in the geologic record. As cells engulfed other cells, some endosymbiotic relationships developed and became permanent. The engulfed cells became organelles within their hosts more than 1 billion years ago, and we see these organelles today as chloroplasts and mitochondria. Loose clusters of unicellular organisms led to cell specialization, and more permanent assemblies produced multicellular organisms. Diversification of body plans became more rapid about 600 million years ago, eventually generating all the major types of organisms we observe today.
10
Evolution by Natural Selection Requires Variation and Competition
Charles Darwin (1809–

The Origin of Species had a tremendous and immediate influence on scientific thought, due in part to the huge volume of work it described and in part to the story it told. Darwin contributed a detailed body of evidence to support a range of interconnected ideas—

Natural selection depends on two characteristics of a population: variation and competition (Figure 1-10). However, the source of a population’s variation eluded Darwin. The genetic program that exists in every organism was unknown to him, as were the mechanisms by which it is handed down from one generation to the next. Darwin was unaware that the work that would eventually reveal these mechanisms had been begun by one of his contemporaries, Gregor Mendel (see Chapter 2). Mendel’s work was little appreciated during his lifetime.

11
Darwin’s ideas have been expanded and developed into a modern synthesis of the theory of evolution, a direct outgrowth of the development of genetics in the early twentieth century—
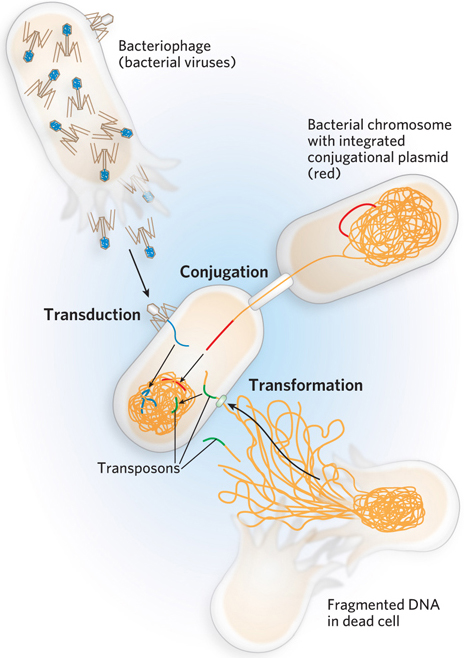
Darwin’s theory of natural selection provides a mechanism that responds directly to the environment. Most of the genetic changes that do not kill an organism produce only small changes in protein function or expression, resulting in a small change in the whole organism. As a result, evolutionary change is usually gradual. Sufficient diversification leads to new species and, with time, to new genera and phyla.
On the most practical level, our connectivity with other species through the tree of life has a critical effect on the study of molecular biology: we can learn about ourselves by studying other organisms (as described in detail in the Model Organisms Appendix). Even the simplest organisms have much to teach us about the inner workings of our own cells. As we will see throughout this book, the processes involved in the flow of biological information, though common to all organisms, are often much more complex in eukaryotes than in bacteria. Much of our understanding of these processes is due to groundbreaking research on bacteria or yeast, followed by further research on more complex model organisms such as worms, insects, or mice (Figure 1-12). In this way, the elucidation of gene functions in a relatively simple organism such as yeast can lead to cures for human disease. Discoveries made in bacteria can generate improvements in agriculture. Fruit flies instruct us about the intricacies of human cognition and the complexities of fetal development. Pandemics of the future can be predicted and tamed by studying the pathogens of the past. Each investigation into the molecular biology of an organism is made more valuable by the fact that all species are related through a shared evolutionary history.
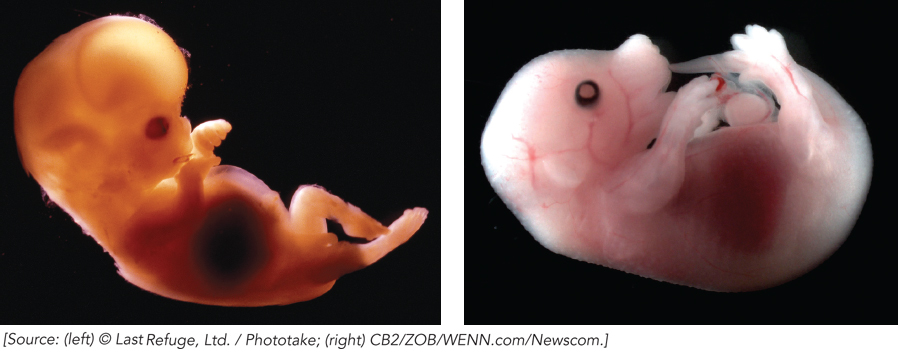
12
As Darwin remarked in The Origin of Species, “There is grandeur in this view of life, with its several powers, having been originally breathed into a few forms or into one; and that, whilst this planet has gone cycling on according to the fixed law of gravity, from so simple a beginning endless forms most beautiful and most wonderful have been, and are being, evolved.”
SECTION 1.1 SUMMARY
Living systems have definable characteristics and requirements. Catalysis and biological information are particularly important requirements for any life form.
The first molecule that fulfilled the requirements of catalysis and biological information may have been a self-
replicating RNA, according to the RNA world hypothesis. LUCA, the last universal common ancestor of all life now present on Earth, can be studied by identifying the common characteristics of living organisms and defining the minimal complement of genes necessary to support a living cell.
Evolution by natural selection is a result of genetic variation within a population and competition between individuals for limited resources. Darwin’s theory of evolution by natural selection has been strengthened by modern studies that reveal the sources of genetic variation, mutation, and recombination.
A common evolutionary heritage links all organisms, allowing the study of model organisms to aid in our understanding of ourselves.