15-2 Cognition and the Association Cortex
Altogether, the primary sensory and motor cortical regions occupy about a third of the neocortex (Figure 15-1). The remaining two-
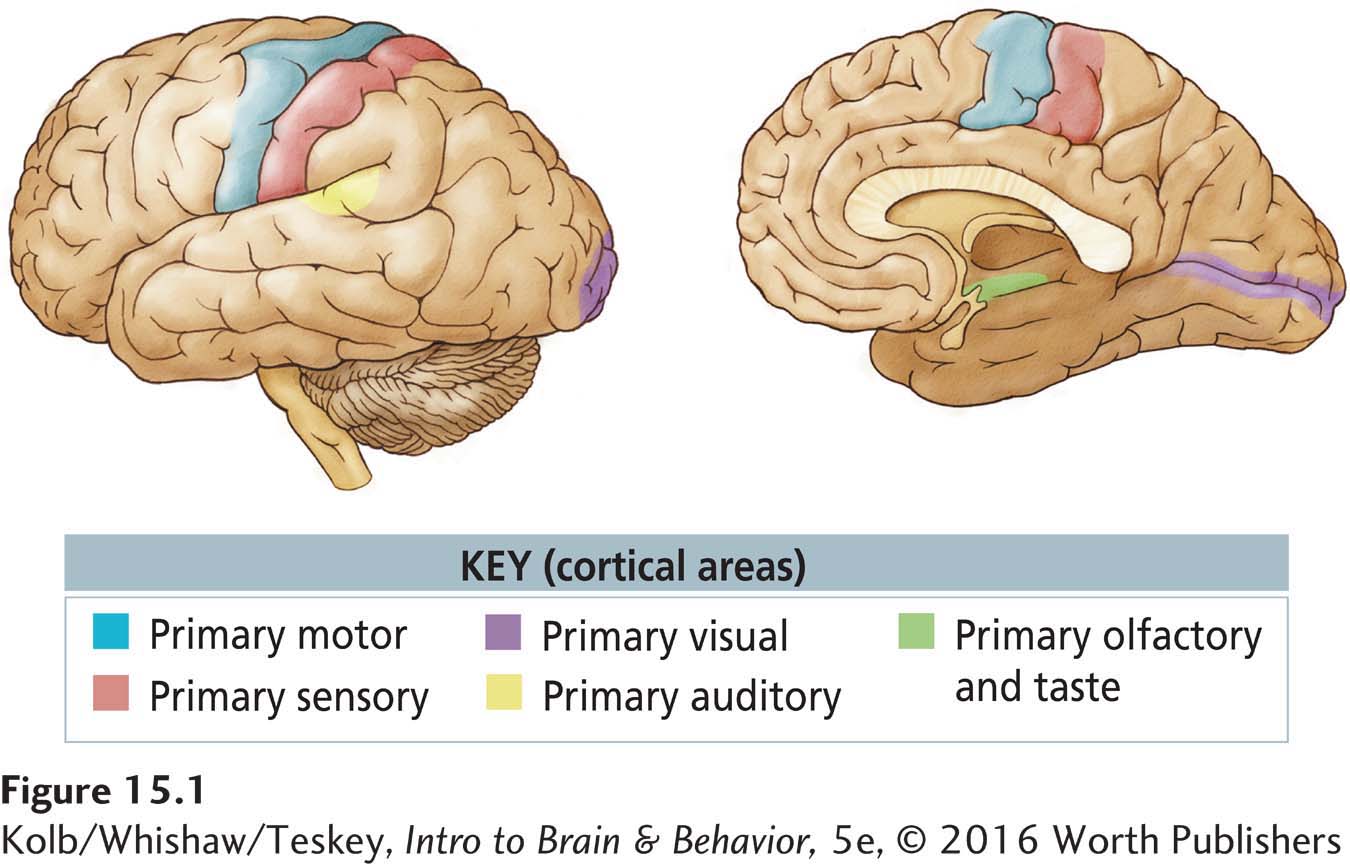
A fundamental difference between association cortex and primary sensory and motor areas is that the association cortex has a distinctive pattern of connections. A major source of input to all cortical areas is the thalamus, which rests atop the brainstem. The primary sensory cortex receives inputs from thalamic nuclei that receive information from the body’s sense organs. But inputs to the association cortex come from thalamic areas that receive their inputs from other cortical regions. As a result, inputs to the association cortex are already highly processed. So this information must be fundamentally different from the raw information reaching the primary sensory and motor cortex. The association regions contain knowledge either about our external or internal world or about movements.
Owing to its close relationship to the visual and auditory sensory regions, the temporal association regions tend to produce cognition related to visual and auditory processing. Similarly, the parietal cortex is closely related to somatosensation and movement control. In contrast, the frontal cortex coordinates information coming from the parietal and temporal association regions with information coming from subcortical regions.
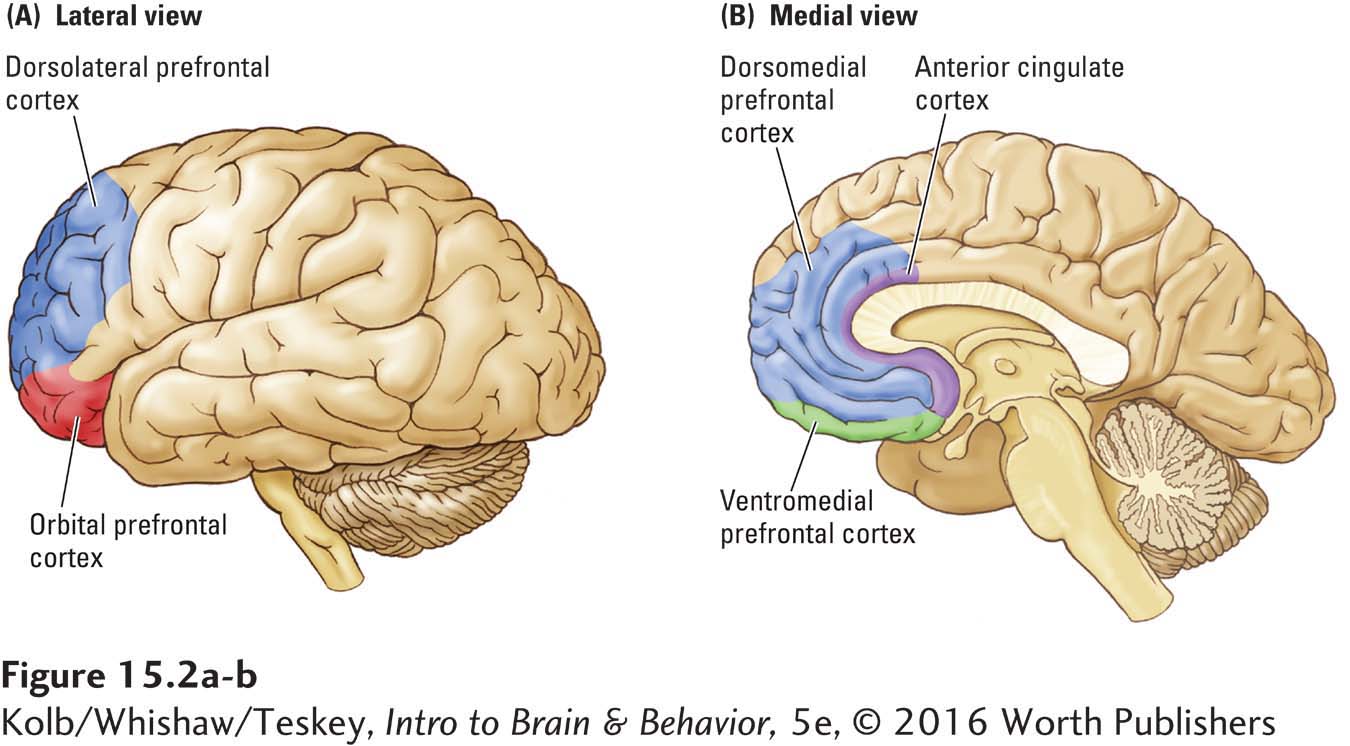
As diagrammed in Figure 15-2, the multiple subdivisions of the prefrontal cortex (PFC) encompass its dorsal, lateral, orbital, and medial regions. Activity in each prefrontal region is associated with the types of cognitive processing that we describe throughout this chapter. An additional frontal lobe region shown in Figure 15-2 is the anterior cingulate cortex (ACC). Although the ACC was once believed only to play a role in emotion, it is becoming clear that it functions as an interface between emotion and cognition.
To understand the types of knowledge that the association areas contain, we next consider discrete cognitive behaviors, then trace these behaviors to different parts of the association cortex.
Knowledge about Objects
Visualize a milk carton sitting on a counter directly in front of you. What do you see? Now imagine moving the carton a few inches off to one side as you continue to stare directly ahead. What do you see now? Next, imagine that you tilt the carton toward you at a 45° angle. Again, what do you see? Probably you answered that you saw the same thing in each situation: a rectangular box with lettering on it.
Intuitively, you feel that the brain must see the object much as you have perceived it. The brain’s seeing, however, is more compartmentalized than are your perceptions. Compartmentalization is revealed in people with damage to various regions of the occipital cortex, many of whom lose one particular aspect of visual perception. For instance, those with damage to visual area V4 can no longer perceive color, whereas those with damage to area V5 can no longer see movement (when the milk carton moves, it becomes invisible to them).
Moreover, your perception of the milk carton’s consistently rectangular shape does not always match the forms your visual system is processing. When you tip the carton toward you, you still perceive it as rectangular, even though it is no longer presenting a rectangular shape to your eyes. Your brain has somehow ignored the change in information about shape that your retinas have sent it and concluded that this object, no matter its apparent shape, remains the same milk carton.
Section 9-4 recounts cases illustrative of various visual-
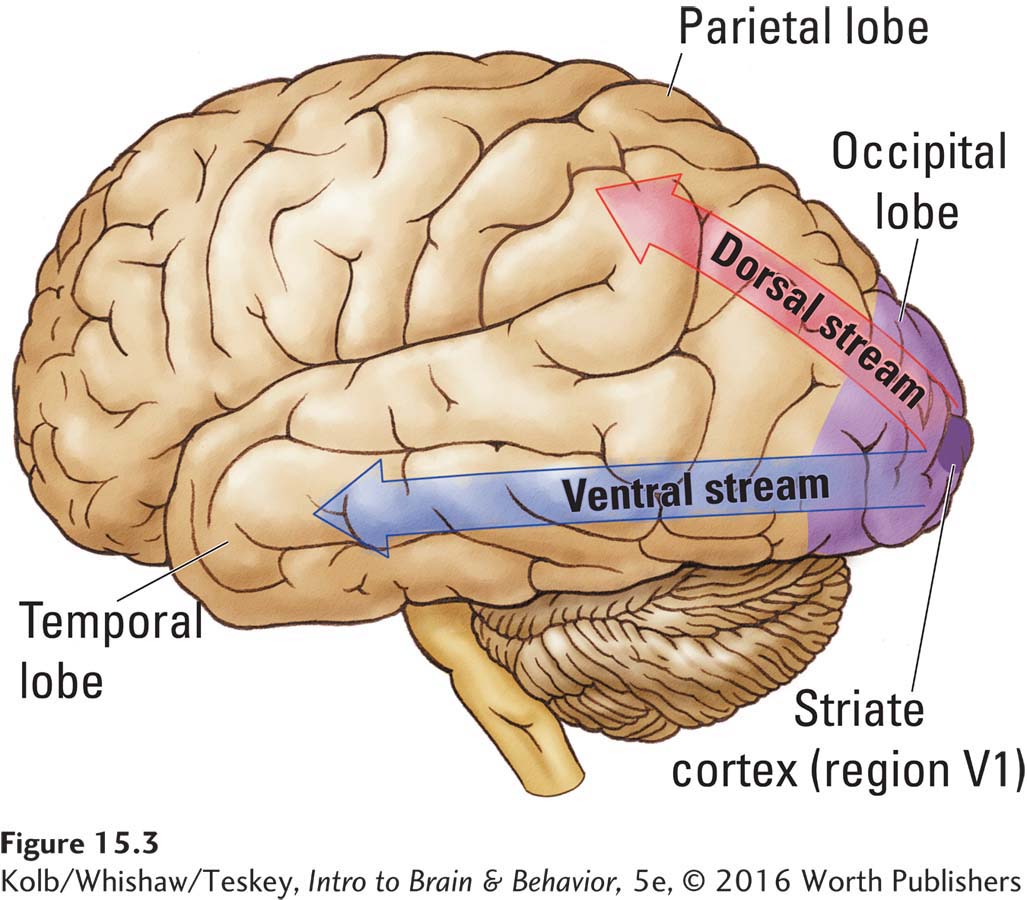
There is more to your conception of the milk carton than merely perceiving and processing its physical characteristics. You also know what a milk carton is, what it contains, and where you can get one. The knowledge about milk cartons that you have acquired is represented in the temporal association cortex, which forms the ventral stream of visual processing. If the temporal association regions are destroyed, a person loses visual knowledge not only about milk cartons but also about all other objects. The person becomes agnosic (unknowing).
Knowledge about objects includes even more than how they look and what they are used for. It depends on what will be done with the information—
Multisensory Integration
Our knowledge about the world comes through multisensory channels. When we see and hear a barking dog, the visual information and auditory information fit together seamlessly. How do all our neural systems and functional levels combine to afford us a unified conscious experience?
Philosophers, impressed with this integrative capacity, identified the binding problem, which asks how the brain ties its single and varied sensory and motor events together into a unified perception or behavior. It is gradually becoming clear how the brain binds up our perceptions and how this ability is gradually acquired in postnatal life (see the review by Stein & Rowland, 2011).
The senses of smell and taste combine to produce the experience of flavor; see Section 12-2.
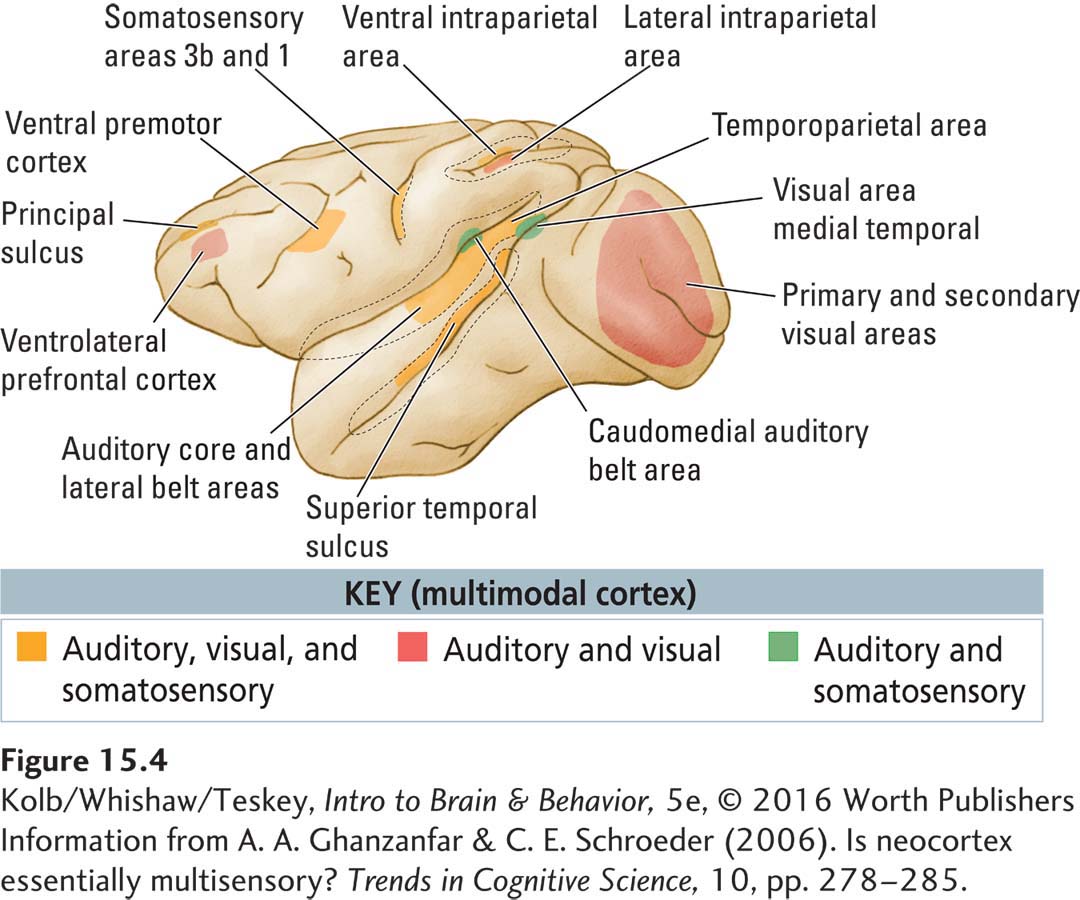
One solution to the sensory integration aspect of the binding problem lies in multimodal regions of the association cortex, that is, regions populated by neurons that respond to information from more than one sensory modality, as illustrated in Figure 15-4. Investigators presume that multimodal regions combine characteristics of stimuli across different senses when we encounter them separately or together. For example, the fact that we can visually identify objects that we have only touched implies a common perceptual system linking the visual and somatic circuits. In Section 15-5, Clinical Focus 15-6 profiles a man in whom stimulation from one sensory modality (taste) concurrently induces the experience of a different modality (touch).
Spatial Cognition
The location of objects is just one aspect of what we know about space. Spatial cognition encompasses a whole range of mental functions that vary from navigational ability (getting from point A to point B) to mentally manipulating complex visual arrays like those shown in Figure 15-5.
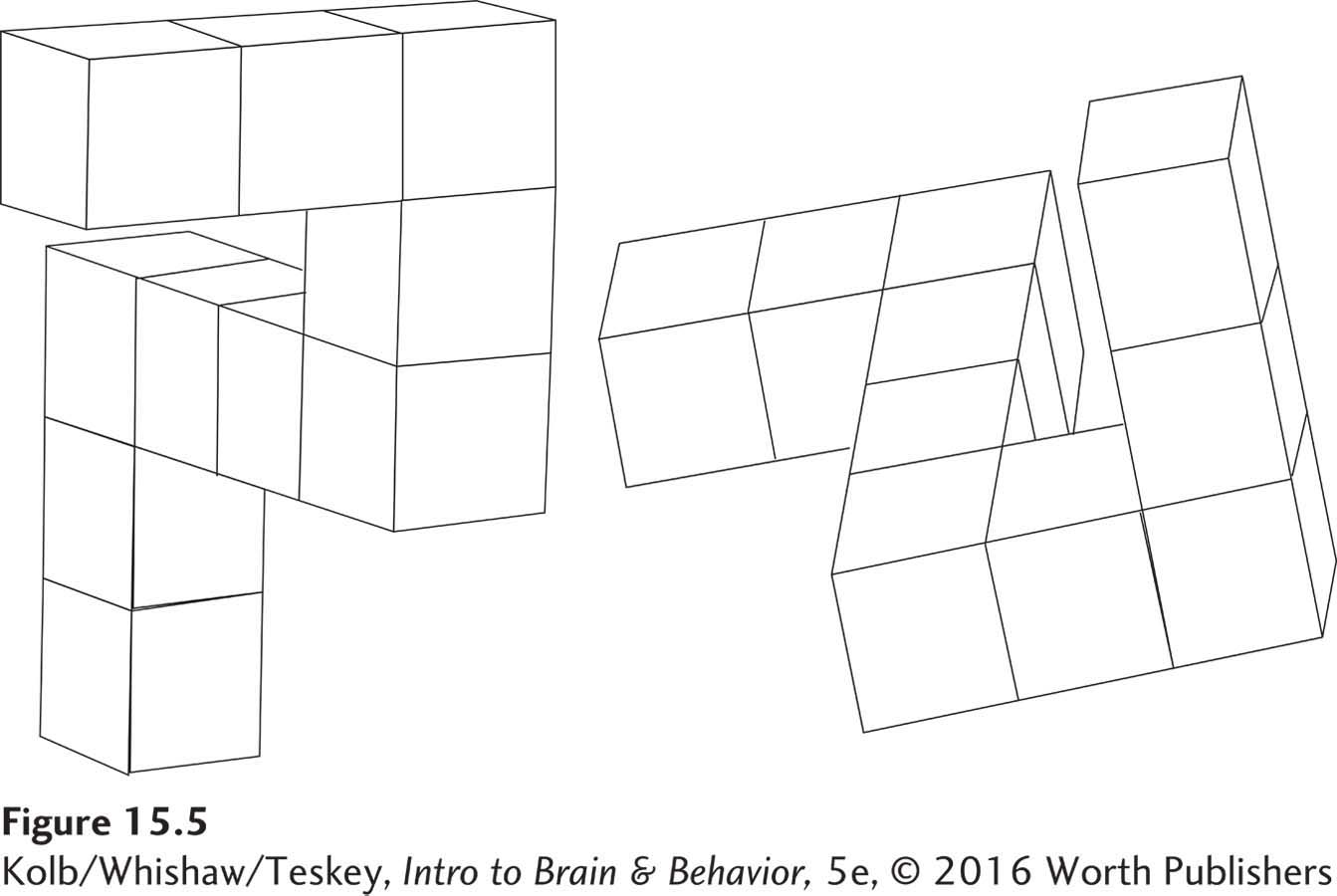
Imagine going for a walk in an unfamiliar park. You do not go around and around in circles. Rather, you proceed in an organized, systematic way. You also need to find your way back. These abilities require a representation of the physical environment in your mind’s eye.
At some time during the walk, let’s assume you are uncertain where you are—
The Basics, Section 1-3, traces nervous system evolution across the animal kingdom.
The evolution of skill in mental manipulation is also closely tied to the evolution of physical movements. It is likely that animals first moved using whole-
Alex the parrot manipulated objects with his beak.
It seems unlikely that more sophisticated cognitive abilities evolved on their own. Why would a fish, say, be able to manipulate an object in its mind that it could not manipulate in the real world? But a human, who can manipulate objects by hand, should be able to imagine such manipulations. After all, we are constantly observing our hands or feet manipulating things: we must have many mental representations of such activities.
Once the brain can manipulate objects that are physically present, it seems a small step to manipulating imagined objects—
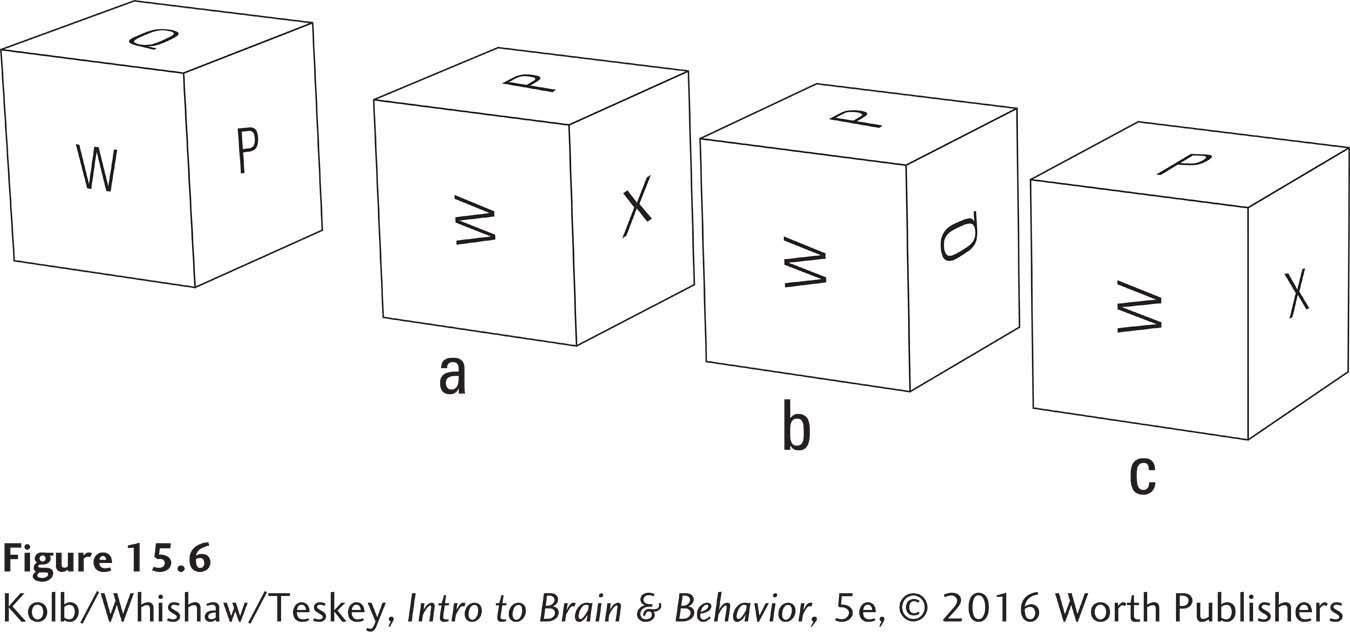
Research findings provide clues to the brain regions participating in various aspects of spatial cognition. For instance, the dorsal stream in the parietal lobes is central in controlling vision for action. Humans make discrete limb movements to points in space, so a reasonable supposition is that the dorsal stream’s evolutionary development provided a neural basis for such spatial cognitive skills as mentally rotating objects. In fact, people with damaged parietal association regions, especially in the right hemisphere, have deficits in processing complex spatial information, both in the real world and in their imagination.
By tracing the evolutionary development of the human brain, we find that the parietal association regions expanded considerably more in humans than in other primates. This expanded brain region functions in part to perform the complex spatial operations just discussed. Humans have a capacity for building that far exceeds that of our nearest relative, the chimpanzee. You may consider it a long leap of logic, but perhaps our increased capacity for building and manipulating objects played heavily into our developing cognitive spatial abilities.
Attention
Imagine you’re meeting some friends at a football game. You search for them as you meander through the crowd in the stadium. Suddenly you hear one friend’s distinctive laugh and turn to scan in that direction. You see your group and rush to join them.
This everyday experience demonstrates the nature of attention, selective narrowing or focusing of awareness to part of the sensory environment or to a class of stimuli. Even as sounds, smells, feelings, and sights bombard you, you still can detect a familiar laugh or spot a familiar face: you can direct your attention.
More than 100 years ago, William James (1890) defined attention: “It is the taking possession by the mind in clear and vivid form of one out of what seem several simultaneous objects or trains of thought.” James’s definition goes beyond our example of locating friends in a crowd, inasmuch as he notes that we can attend selectively to thoughts as well as to sensory stimuli. Who hasn’t at some time been so preoccupied with a thought as to exclude all else from mind? So attention can be directed inward as well as outward.
Selective Attention
The answer to the mental manipulation in Figure 15-6 is a.
As with many other inferred mental processes, studying the neural basis of attention is challenging. Research with monkeys trained to attend to particular locations or visual stimuli, however, has identified neurons in the cortex and midbrain that show enhanced firing rates to particular locations or visual stimuli. Significantly, the same stimulus can activate a neuron at one time but not at another, depending on the monkey’s learned focus of attention.
In the study shown in Experiment 15-2, James Moran and Robert Desimone (1985) trained monkeys to hold a bar while gazing at a fixation point on a screen. A sample stimulus (e.g., a vertical red bar) appeared briefly at one location in the visual field, followed about 500 milliseconds later by a test stimulus at the same location. When the test stimulus was identical with the initial sample stimulus, an animal was rewarded if it immediately released the bar.
EXPERIMENT
Question: Can neurons learn to respond selectively to stimuli?
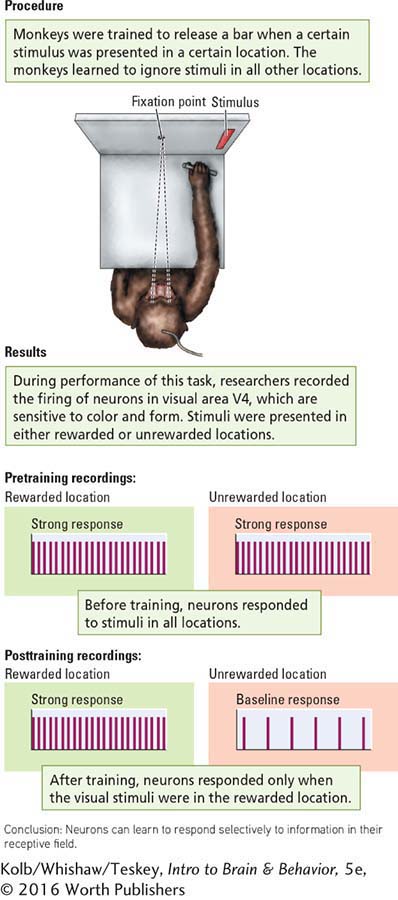
Each animal was trained to attend to stimuli presented in one particular area of the visual field and to ignore stimuli in any other area. In this way, the same visual stimulus could be presented to different regions of a neuron’s receptive field to test whether the cell’s response varied with stimulus location.
As the animals performed the task, the researchers recorded neurons firing in visual area V4. Neurons in V4 are sensitive to color and form, and different neurons respond to different combinations of the two variables (e.g,, a red vertical bar or a green horizontal bar). Visual stimuli were presented either in the correct location for a reward or in an incorrect location for no reward.
As diagrammed in the Results section of Experiment 15-2, neurons responded only when a visual stimulus was in the reward location, even though the same stimulus was presented in a no-
Deficits of Attention
Attention is probably a property of neurons throughout the brain, with some regions playing more prominent roles than others. The frontal lobes, for instance, are central in attention. People with frontal lobe injuries tend to become overly focused on environmental stimuli. They seem to selectively direct attention to an excessive degree or to have difficulty shifting attention. Studies of these people suggest that the frontal association cortex controls the ability to direct attention flexibly to where it is needed. Indeed, planning, a key frontal lobe function, requires this ability.
That the parietal association cortex is key in other aspects of attention is perhaps best illustrated by the attention deficit referred to as neglect. Neglect occurs when a brain-
People with damage to the parietal association cortex of the right hemisphere may have particularly severe neglect of objects or events in the left side of their world. For example, one man dressed only the right side of his body, shaved only the right side of his face, and read only the right side of a page (if you can call that reading). He could move his left limbs spontaneously, but when asked to raise both arms, he would raise only the right. When pressed, he could be induced to raise the left arm, but he quickly dropped it to his side again.
As people with contralateral neglect begin to recover, they show another interesting symptom. They neglect information on one side of the body when it is simultaneously presented with similar information on the other side of the body. Figure 15-7 shows a common clinical test for this symptom, called extinction.
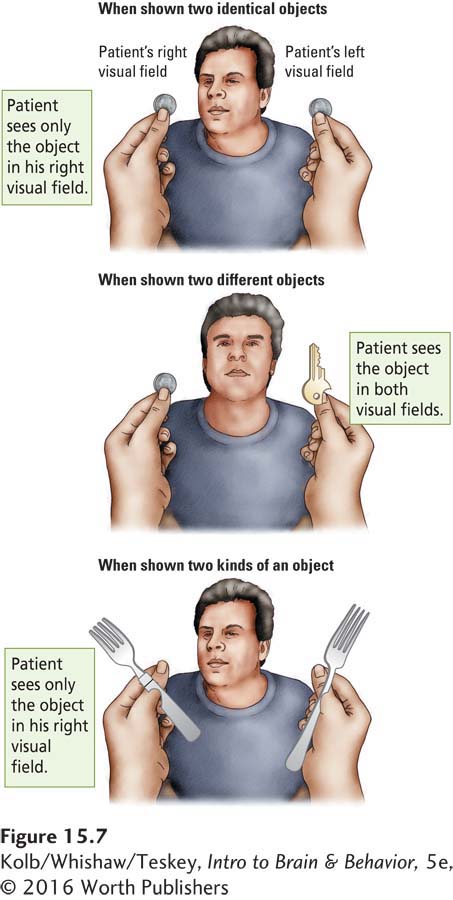
In an extinction test, the patient is asked to keep his or her eyes fixed on the examiner’s face and to report objects presented in one or both sides of the visual field. When presented with a single object (a fork) to one side or the other, the patient orients himself or herself toward the appropriate side of the visual field, so we know that he or she cannot be blind on either side. But now suppose that two forks are presented, one on the left and one on the right. The patient ignores the fork on the left and reports the one on the right. When asked about the left side, the patient is quite certain that nothing appeared there and that only one fork was presented, on the right.
Perhaps the most curious aspect of neglect is that people who have it fail to pay attention not only to one side of the physical world around them but also to one side of the world represented in their mind. We studied one woman who had complete neglect for everything on her left side. She complained that she could not use her kitchen because she could never remember the location of anything on her left.
We asked her to imagine standing at the kitchen door and to describe what was in the various drawers on her right and left. She could not recall anything on her left. We then asked her to imagine walking to the end of the kitchen and turning around. We again asked her what was on her right, the side of the kitchen that had previously been on her left. She broke into a big smile and tears ran down her face as she realized that she now knew what was on that side of the room. All she had to do was reorient her body in her mind’s eye. She later wrote and thanked us for changing her life, because she was now able to cook again. Clearly, neglect can exist in the mind as well as in the physical world.
Although complete contralateral neglect is usually associated with parietal lobe injury, specific forms of neglect can arise from other injuries. Ralph Adolphs and his colleagues (2005) describe the case of S. M., a woman with bilateral amygdala damage who could not recognize fear in faces. On further study, the reason was discovered: S. M. failed to look at the eyes when she looked at faces; instead, she looked at other facial features such as the nose. Because fear is most clearly identified in the eyes, not the nose, she did not identify the emotion. When she was specifically instructed to look at the eyes, her recognition of fear became entirely normal. Thus, the amygdala plays a role in directing attention to the eyes to identify facial expressions.
Planning
At noon on a Friday, a friend proposes that she and you go to a nearby city for the weekend to attend a concert. She will pick you up at 6:00 p.m. and you will drive there together.
Because you are completely unprepared for this invitation and because you are going to be busy until 4:00, you must rush home and get organized. En route you stop at a fast food restaurant so that you won’t be hungry on the 2-
Although the task of getting ready in a hurry may make us a bit harried, most of us can manage it. People with frontal lobe injury cannot. To learn why, let’s consider what the task requires.
To plan your behavior, you must select from many options. What do you need to take with you? Cash? Then which ATM is closest, and what is the quickest route to it? Are you hungry? Then what is the fastest way to get food on a Friday afternoon?
In view of your time constraint, you have to ignore irrelevant stimuli. If you pass a sign advertising a sale in your favorite store, for instance, you have to ignore it and persist with the task at hand.
You have to keep track of what you have done already, a requirement especially important while you are packing. You do not want to forget anything or pack duplicates. You do not want to take four pairs of shoes but no toothbrush.
The task’s general requirements can be described as the temporal (time) organization of behavior. You are planning what you need to do and when you need to do it. Temporal planning is the general function of the frontal lobes, especially the prefrontal cortex.
But to plan, you also need to recognize objects (an occipital and temporal lobe function) and to make appropriate movements with respect to them (a parietal lobe function). You can therefore think of the frontal lobes as acting like an orchestra conductor. The frontal lobes make and read a motor plan to organize behavior in space and time—
Performance on the Wisconsin Card Sorting Test exemplifies the deficits frontal lobe injury causes. Figure 15-8 shows the testing materials. The subject is presented with four stimulus cards bearing designs that differ in color, form, and number of elements, thus setting up three possible sorting categories. The subject must sort a deck of cards into piles in front of the various stimulus cards, depending on the sorting category called for. But the correct sorting category is never stated. The subject is simply told after placing each card whether the choice is correct or incorrect.

In one trial, for example, the first correct sorting category is color. After the subject has sorted a number of cards by color, the correct solution switches, without warning, to form. When the subject has started to sort by form, the correct solution again changes unexpectedly, this time to the number of items on each card. The sorting rule later becomes color again, and so on, with each change in rule coming unannounced.
Shifting response strategies is particularly difficult for people with frontal lobe lesions: they may continue responding to the original stimulus (color) for as many as 100 cards until the test ends. This pattern, known as perseveration, is the tendency to emit repeatedly the same verbal or motor response to varied stimuli.
Frontal lobe subjects may even comment that they know that color is no longer the correct category, but they continue to sort by color. One stated: “Form is probably the correct solution now so this [sorting by color] will be wrong, and this will be wrong, and wrong again.” Despite knowing the correct sorting category, the frontal lobe patient cannot shift behavior in response to the new external information.
Imitation and Understanding
In all communication—
How do both sender and receiver of a potentially ambiguous gesture, such as a raised hand or a faint smile, achieve a common understanding? Giacomo Rizzolatti and his colleagues (Rizzolatti, 2007; Rizzolatti & Craighero, 2004) proposed an answer. In monkeys’ frontal lobes they identified neurons that discharge during active movements of the hand or mouth or both. These neural discharges do not precede the movements but instead occur in synchrony with them. But it takes time for a neural message to go from a frontal lobe to a hand, so we would predict that, if these cells are controlling the movements, they will discharge before the movements take place. The cells must therefore be recording a movement that is taking place.
In the course of his studies, Rizzolatti also found that many “movement” neurons located in the inferior frontal and posterior parietal cortex discharge when a monkey sees other monkeys make the same movements. They also discharge when the monkey sees the experimenter make the movements. Rizzolatti called them mirror neurons. The researchers proposed that mirror neurons represent actions, one’s own or those of others. Such neural representations could be used both for imitating others’ actions and for understanding their meanings, thus enabling appropriate responses. Mirror neurons therefore could provide the link between the sender and the receiver of a communication.
Rizzolatti and his colleagues used PET to look for these same neuron populations in humans. Participants were asked to watch a movement, to make the same movement, or to imagine the movement. Each condition activated a region of the lateral frontal lobe in the left hemisphere, including Broca’s area. Taken together with the results of the monkey studies, this finding suggests that primates have a fundamental mechanism for recognizing action. People apparently recognize others’ actions, because the neural patterns produced when they observe those actions are similar to those produced when they themselves make those same actions. Research Focus 15-3, The Rise and Fall of Mirror Neurons, frames the debate on mirror neuron function.
15-3
The Rise and Fall of Mirror Neurons
Rizzolatti and colleagues’ discovery of mirror neurons in the 1990s was serendipitous (e.g., Rizzolatti, 2007). The group was investigating how neurons in the premotor cortex of macaque monkeys control grasping actions. The monkeys were trained to make different grasping movements to retrieve bits of food. When the animals reached for the food, cells in the premotor cortex were highly active, revealing their role in movement control.
Between reaching trials the monkeys could observe an investigator making reaching movements similar to those the monkey had made. Unexpectedly, many of the same neurons fired in response to the monkeys observing the experimenter’s reaching: the cells appeared to mirror others’ behavior. The name mirror neurons stuck. Studies using noninvasive imaging demonstrated similar neuronal populations, although apparently more widespread, in humans.
Over the ensuing decade, mirror neurons were hypothesized to be the basis of understanding actions rather than merely recognizing them. This possibility had profound implications and led to propositions that mirror neurons were the basis of speech and language, empathy, social cognition, theory of mind—
The lure of mirror neurons spread well beyond neuroscience and into the popular media. All the hype resulted in mirror neurons being widely perceived as among the most important discoveries in neuroscience in the past generation.
But by the early 2000s, cracks began to show in the mirror. Gregory Hickok (2014) has argued that mirror neurons do not make actions understandable, but they do refine movement control. Hickok and others suggest that mirror neurons do not “understand” action, but rather their function arises from an association between executing an action and the self-
Thus, the neurons learn to mirror. But why? What is the benefit? One possibility is that because the actions of others are relevant to one’s own actions, having a system to link others’ perceived actions with one’s own appropriate actions is adaptive. An extension of this idea is that mirror neurons are activated after an action is understood by other brain regions as a way to make predictions about future actions (Hickok, 2013). In this view, mirror neurons simply reflect the fact that the brain understands actions (Csibra, 2007). The advantage of this type of mirror system is that with this knowledge we can make movements faster and more accurately.
The lessons from the rise and fall of mirror neurons: First, although the mirror neuron theory of functioning to understand action may be wrong, it has served the field well by stimulating new research. That research has led to better understanding of how the brain thinks. A second lesson is vigilance: some scientists and media can push scientific theories well beyond the data.
15-2 REVIEW
Cognition and the Association Cortex
Before you continue, check your understanding.
Question 1
The association cortex contains ____________ and functions to produce ____________.
Question 2
As a general rule, the ____________ lobes generate knowledge about objects, whereas the ____________ lobes produce various forms of spatial cognition.
Question 3
The frontal lobes function not only to make movements but also to ____________ and to ____________.
Question 4
____________ neurons in the frontal and parietal lobes represent actions, one’s own or those of others.
Question 5
Describe the function of multimodal cortex.
Answers appear in the Self Test section of the book.