Localization of mRNAs Permits Production of Proteins at Specific Regions Within the Cytoplasm
Many cellular processes depend on localization of particular proteins to specific structures or regions of the cell. In later chapters, we examine how some proteins are transported after their synthesis to their proper cellular location. Alternatively, protein localization can be achieved by localization of mRNAs to the specific regions of the cytoplasm in which their encoded proteins function. In most cases examined thus far, such mRNA localization is specified by sequences in the 3′ UTR of the mRNA. A recent genomic-level study of mRNA localization in Drosophila embryos revealed that some 70 percent of the 3000 mRNAs analyzed were localized to specific subcellular regions, raising the possibility that mRNA localization is a much more general phenomenon than previously appreciated.
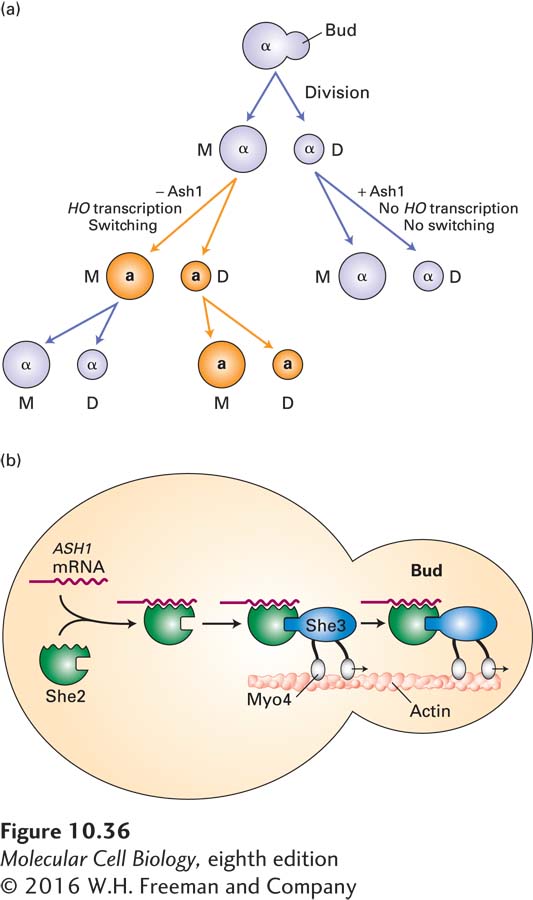
Localization of mRNAs to the Bud in S. cerevisiae The most thoroughly understood example of mRNA localization occurs in the budding yeast S. cerevisiae. As discussed in Chapter 9, whether a haploid yeast cell exhibits the a or α mating type is determined by whether a or α genes are present at the expressed MAT locus on chromosome III (see Figure 9-35). The process that transfers a or α genes from the silent mating-type locus to the expressed MAT locus is initiated by a sequence-specific endonuclease called HO. Transcription of the HO gene is dependent on the SWI/SNF chromatin-remodeling complex (see Section 9.5). Daughter yeast cells that arise by budding from mother cells contain a transcriptional repressor called Ash1 (for Asymmetric synthesis of HO) that prevents recruitment of the SWI/SNF complex to the HO gene, thereby preventing its transcription. The absence of Ash1 from mother cells allows them to transcribe the HO gene. As a consequence, mother cells switch their mating type, while daughter cells generated by budding do not (Figure 10-36a).
FIGURE 10-36Switching of mating type in haploid yeast cells. (a) Division by budding forms a larger mother cell (M) and smaller daughter cell (D), both of which have the same mating type as the original cell (α in this example). The mother cell can switch mating type during G1 of the next cell cycle and then divide again, producing two cells of the opposite type (a in this example). Switching depends on transcription of the HO gene, which occurs only in the absence of Ash1 protein. The smaller daughter cells, which produce Ash1 protein, cannot switch; after growing in size through interphase, they divide to form a mother cell and daughter cell. (b) Model for restriction of mating-type switching to mother cells in S. cerevisiae. Ash1 protein prevents a cell from transcribing the HO gene, whose encoded protein initiates the DNA rearrangement that results in mating-type switching from a to α or α to a. Switching occurs only in the mother cell, after it separates from a newly budded daughter cell, because the Ash1 protein is present only in the daughter cell. The molecular basis for this differential localization of Ash1 is the one-way transport of ASH1 mRNA into the bud. A linking protein, She2, binds to specific 3′ untranslated sequences in the ASH1 mRNA and also binds to She3 protein. This protein, in turn, binds to a myosin motor, Myo4, which moves along actin filaments into the bud. See S. Koon and B. J. Schnapp, 2001, Curr. Biol.11:R166.
Ash1 protein accumulates only in daughter cells because the mRNA encoding it is localized to daughter cells. The localization process requires three proteins: She2 (for SWI-dependent HO expression), an RNA-binding protein that binds specifically to a localization signal with a specific RNA structure in the ASH1 mRNA; Myo4, a myosin motor protein that moves cargoes along actin filaments (see Chapter 17); and She3, which links She2, and therefore ASH1 mRNA, to Myo4 (Figure 10-36b). ASH1 mRNA is transcribed in the nucleus of the mother cell before mitosis. Movement of Myo4, with its bound ASH1 mRNA, along actin filaments that extend from the mother cell into the bud carries the ASH1 mRNA into the growing bud before cell division.
Page 458
Page 459
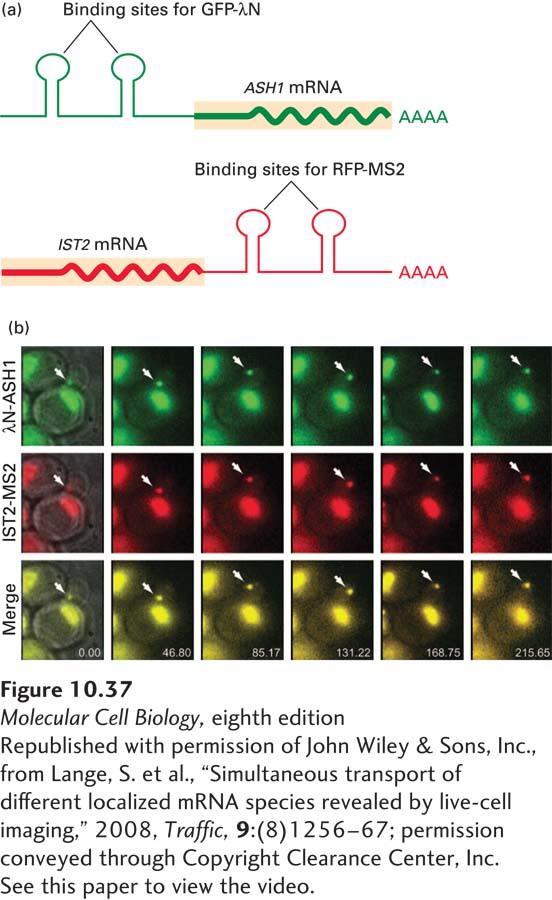
At least 23 other mRNAs were found to be transported by the She2/She3/Myo4 system. All have an RNA localization signal to which She2 binds, usually in the 3′ UTR. The transport process can be visualized in live cells by the experiment shown in Figure 10-37. RNAs can be fluorescently labeled by including in their sequence high-affinity binding sites for RNA-binding proteins, such as bacteriophage MS2 coat protein and bacteriophage λ N protein, which bind to different stem-loops with specific sequences (Figure 10-37a). When such engineered mRNAs are expressed in budding yeast cells, along with the bacteriophage proteins fused to proteins that fluoresce different colors, the fusion proteins bind to their specific RNA binding sites, thereby labeling the RNAs that contain those sites with different colors. In the experiment shown in Figure 10-37b, ASH1 mRNA was labeled by the binding of green fluorescent protein fused to λN. Another mRNA localized to the bud by the same transport system, the IST2 mRNA, which encodes a component of the growing bud membrane, was labeled by the binding of red fluorescent protein fused to MS2 coat protein. Video of a budding cell showed that the differently labeled ASH1 and IST2 mRNAs accumulated in the same large cytoplasmic RNP particle, containing multiple mRNAs, in the mother-cell cytoplasm, as can be seen from the merge of the green and red fluorescent signals. The RNP particle was then transported into the bud within about one minute.
EXPERIMENTAL FIGURE 10-37Transport of mRNP particles from a yeast mother cell into the bud. (a) Yeast cells were engineered to express an ASH1 mRNA with binding sites for the bacteriophage λ N protein in its 5′ UTR, and an IST2 mRNA with binding sites for bacteriophage MS2 coat protein in its 3′ UTR. A fusion of green fluorescent protein to λ N protein (GFP-λN) and a fusion of red fluorescent protein to MS2 coat protein (RFP-MS2) were also expressed in the same cells. In other experiments, these fluorescently tagged sequence-specific RNA-binding proteins were shown to bind to their own specific binding sites engineered into the ASH1 and IST2 mRNAs, and not to each other’s binding sites. Both fluorescently tagged bacteriophage proteins also contained a nuclear localization signal so that those proteins that were not bound to their high-affinity binding sites in these mRNAs were transported into nuclei through nuclear pore complexes (see Chapter 13). This step was necessary to prevent high fluorescence from excess GFP-λN and RFP-MS2 in the cytoplasm. (b) Frames from a video of fluorescing cells. GFP-λN and RFP-MS2 were independently visualized by using millisecond alternating laser excitation of GFP and RFP. The nucleus next to the large vacuole in the mother cell near the center of each micrograph, as well as nuclei in neighboring cells, was observed by green and red fluorescence, as shown in the top and middle rows. A merge of the two images is shown in the bottom row, which also indicates the time elapsed between images. An RNP particle containing both the ASH1 mRNA with λN-binding sites and the IST2 mRNA with MS2-binding sites was observed in the mother-cell cytoplasm in the left column of images (arrow). The particle increased in intensity between 0.00 and 46.80 seconds, indicating that more of these mRNAs joined the RNP particle. The RNP particle was transported into the bud between 46.80 and 85.17 seconds and then became localized to the bud tip.
[Republished with permission of John Wiley & Sons, Inc., from Lange, S. et al., “Simultaneous transport of different localized mRNA species revealed by live-cell imaging,” 2008, Traffic,9:(8)1256–67; permission conveyed through Copyright Clearance Center, Inc. See this paper to view the video.]
Formation of large cytoplasmic RNP particles like those observed in Figure 10-37b, and in other examples of transported RNA in cells of higher eukaryotes, requires low-complexity amino acid sequences, such as sequences composed of repeats of [G/S]Y[G/S], in the RNA-binding protein. Peptides containing these low-complexity sequences spontaneously associate in vitro, forming a semi-permeable gel. These gels can be dissociated by phosphorylation of serines within them. Such complexes are probably involved in the formation of the large RNP complexes transported on actin cables in yeast and on microtubules in large asymmetric cells in higher eukaryotes (see Chapter 17 and 18), such as the neurons described in the next section. Regulated phosphorylation of these low-complexity sequences in RNA-binding proteins associated with RNP particles may well account for the regulated formation and dissociation of RNP particles such as those observed in Figure 10-37b.
Localization of mRNAs to Synapses in the Mammalian Nervous System As mentioned earlier, localization of specific mRNAs at synapses far from the nucleus of a neuron plays an essential role in learning and memory (Figure 10-38). Like the localized mRNAs in yeast, these mRNAs contain RNA localization signals in their 3′ UTR. Some of these mRNAs are initially synthesized with short poly(A) tails that do not allow translation initiation. Once again, large RNP particles containing multiple mRNAs bearing localization signals form in the cytoplasm near the nucleus. In this case, the RNP particles are then transported down the axon to synapses by kinesin motor proteins, which travel down microtubules extending the length of the axon (see Chapter 18). Electrical activity at a given synapse may then stimulate polyadenylation of the mRNAs in the region of that synapse, activating the translation of encoded proteins that increase the size and alter the neurophysiological properties of that synapse, while leaving unaffected the hundreds to thousands of other synapses made by the neuron.
Page 460
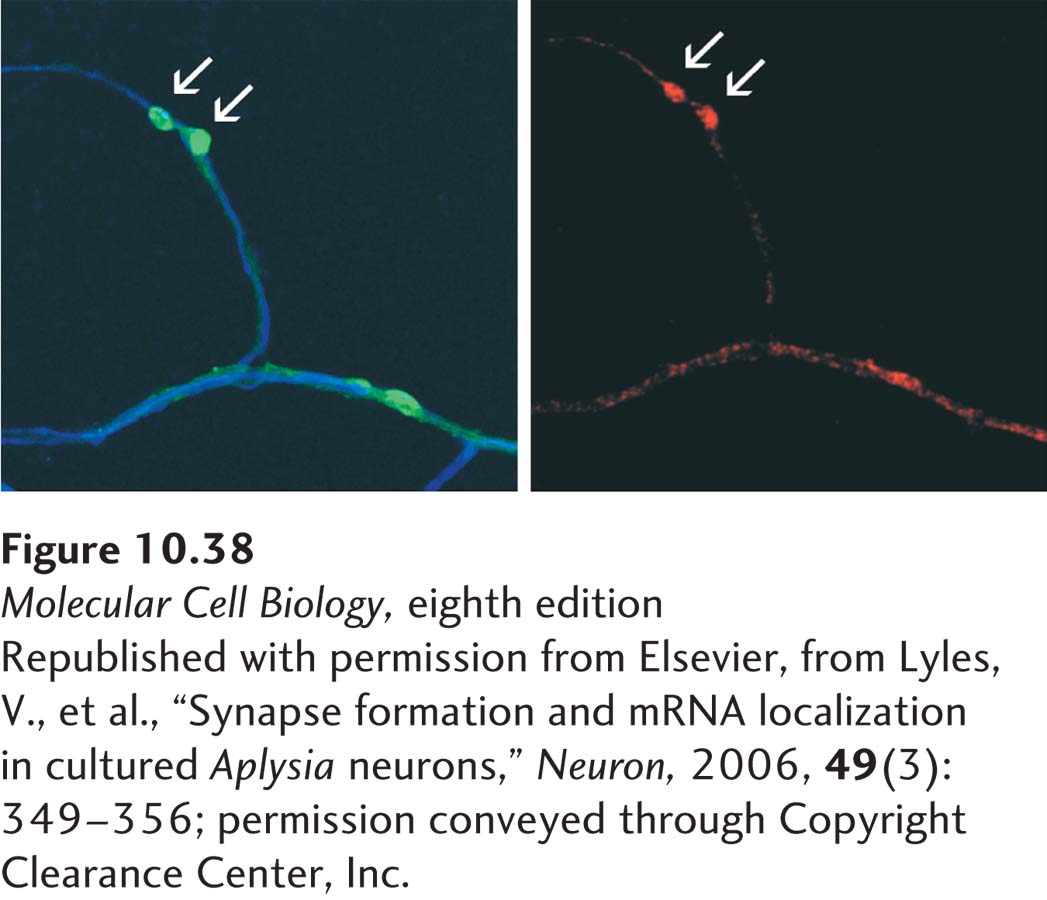
EXPERIMENTAL FIGURE 10-38A specific neuronal mRNA localizes to synapses. Sensory neurons from the sea slug Aplysia californica were cultured with target motor neurons so that processes from the sensory neurons formed synapses with processes from the motor neurons. The micrograph at the left shows motor neuron processes visualized with a blue fluorescent dye. GFP-VAMP (green) was expressed in sensory neurons and marks the location of synapses formed between sensory and motor neuron processes (arrows). The micrograph at the right shows red fluorescence from in situ hybridization of an antisensorin mRNA probe. Sensorin is a neurotransmitter expressed by the sensory neuron only; sensory neuron processes are not otherwise visualized in this preparation, but they lie adjacent to the motor neuron processes. The in situ hybridization results indicate that sensorin mRNA is localized to synapses.
[Republished with permission from Elsevier, from Lyles, V., et al., “Synapse formation and mRNA localization in cultured Aplysia neurons,” Neuron, 2006, 49(3):349–356; permission conveyed through Copyright Clearance Center, Inc.]