Mitochondria Are Dynamic Organelles That Interact Directly with One Another
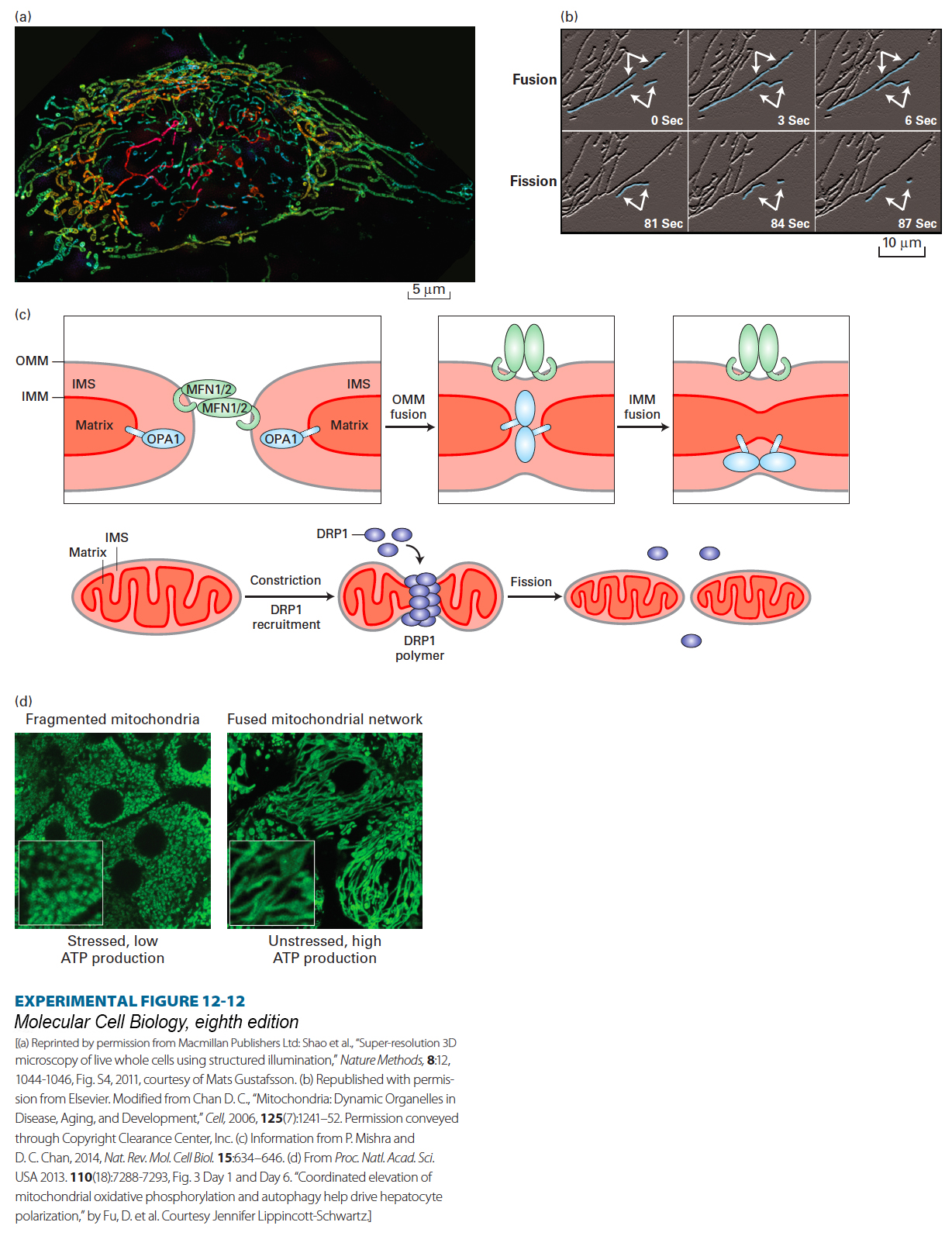
Analysis of fluorescently labeled mitochondria in live cells has shown that mitochondria in many different types of cells are highly dynamic. They undergo frequent fusions (merging) and fissions (breaking apart) that generate tubular, sometimes branched networks (Figures 12-12a and b), which may account for some of the wide variety of mitochondrial morphologies seen in different types of cells. When individual mitochondria fuse, each of the two membranes fuses (inner with inner, and outer with outer) and each of their distinct compartments intermix (matrix with matrix, intermembrane space with intermembrane space). A set of four evolutionarily conserved GTP-hydrolyzing (GTPase) enzymes—MFN1 (mitofusin 1), MFN2 (mitofusin 2), OPA1, and DRP1—plays critical roles in mediating these membrane fusions and fissions (Figure 12-12c). These enzymes are members of the dynamin family of GTPases. Dynamin, the first-identified member of this family, mediates a comparable membrane fission reaction required during the pinching off of endocytic vesicles from the plasma membrane (see Chapter 14). Mutations in several of the genes encoding these GTPases can disrupt mitochondrial functions, such as maintenance of proper inner membrane electric potential, and structure, resulting in truncated or enlarged mitochondria, and cause human disease. The inherited, autosomal dominant neuromuscular disease Charcot-Marie-Tooth subtype 2A is caused by loss-of-function mutations in MFN2 that lead to defects in peripheral nerve function and progressive muscle weakness, mainly in the feet and hands. Mutations in OPA1 are associated with autosomal dominant optic atrophy that influences nerves in the eye’s retina.
[(a) Reprinted by permission from Macmillan Publishers Ltd: Shao et al., “Super-resolution 3D microscopy of live whole cells using structured illumination,” Nature Methods,8:12, 1044-1046, Fig. S4, 2011, courtesy of Mats Gustafsson. (b) Republished with permission from Elsevier. Modified from Chan D. C., “Mitochondria: Dynamic Organelles in Disease, Aging, and Development,” Cell, 2006, 125(7):1241–52. Permission conveyed through Copyright Clearance Center, Inc. (c) Information from P. Mishra and D. C. Chan, 2014, Nat. Rev. Mol. Cell Biol.15:634–646. (d) From Proc. Natl. Acad. Sci. USA 2013. 110(18):7288-7293, Fig. 3 Day 1 and Day 6. “Coordinated elevation of mitochondrial oxidative phosphorylation and autophagy help drive hepatocyte polarization,” by Fu, D. et al. Courtesy Jennifer Lippincott-Schwartz.]
EXPERIMENTAL FIGURE 12-12Mitochondria undergo rapid fusion and fission. (a) A human HeLa cell labeled with a mitochondrion-specific fluorescent dye (MitoTracker Green) was imaged using three-dimensional structured illumination fluorescence microscopy (a 6.1-µm-thick section through the cell is shown). The network of fused and branched mitochondria is seen in the cytoplasm, with only a few mitochondria observed above or below the nucleus (unstained central dark oval). The identity of the striations seen within the mitochondria is not known. The mitochondria are shown in artificial colors to indicate their positions relative to the surface to which the cell is attached (blue is closest to and red farthest from the surface). (b) Mitochondria labeled with a fluorescent protein in a live normal mouse embryonic fibroblast were observed using time-lapse fluorescence microscopy. Several mitochondria undergoing fusion (top) or fission (bottom) are artificially highlighted in blue and with arrows. (c) Mitochondrial fusion (top) and fission (bottom) are mediated by a set of GTPase enzymes (MFN1, MFN2, OPA1, and DRP1). The integral membrane proteins MFN1 and MFN2 (MFN1/2) mediate outer mitochondrial membrane (OMM) fusion, which is followed by fusion of the inner mitochondrial membranes (IMM) mediated by the integral membrane protein OPA1. The matrix and inner membrane space (IMS) remain distinct. The soluble cytosolic GTPase DRP1 is recruited to a constricted site on the surface of a mitochondrion, where DRP1 polymers sever the membrane, resulting in fission. A variety of post-translational modifications of DRP1 regulate fission. (d) (Left) Rat liver cells (hepatocytes) one day after being removed from the liver and placed in cell culture, are stressed and depolarized (lack some of the morphological and biochemical properties of epithelial cells; see Chapter 20), have low levels of oxidative phosphorylation and ATP production, and have fragmented mitochondria (visualized by staining with MitoTracker Green). (Right) After growth in culture for six days, the hepatocytes become polarized, their mitochondria fuse, forming an extensive network, and the cells exhibit high levels of oxidative phosphorylation and ATP production. Insets show higher-magnification views of the mitochondria.
[(a) Reprinted by permission from Macmillan Publishers Ltd: Shao et al., “Super-resolution 3D microscopy of live whole cells using structured illumination,” Nature Methods,8:12, 1044-1046, Fig. S4, 2011, courtesy of Mats Gustafsson. (b) Republished with permission from Elsevier. Modified from Chan D. C., “Mitochondria: Dynamic Organelles in Disease, Aging, and Development,” Cell, 2006, 125(7):1241–52. Permission conveyed through Copyright Clearance Center, Inc. (c) Information from P. Mishra and D. C. Chan, 2014, Nat. Rev. Mol. Cell Biol.15:634–646. (d) From Proc. Natl. Acad. Sci. USA 2013. 110(18):7288-7293, Fig. 3 Day 1 and Day 6. “Coordinated elevation of mitochondrial oxidative phosphorylation and autophagy help drive hepatocyte polarization,” by Fu, D. et al. Courtesy Jennifer Lippincott-Schwartz.]
Page 529
What is the value of mitochondrial fission and fusion? It has been suggested that fusion helps to maintain a relatively homogeneous population of mitochondria within a cell. Should some subset of individual mitochondria suffer deleterious modification or loss of important components, fusion with other mitochondria would permit restoration by sharing of those components. Studies of cells and organisms with mutations in genes encoding the fusion machinery suggest that fusion also plays a role in the proper localization of mitochondria within cells, maintaining the proper morphology and cristal organization, distribution of mitochondrial DNA, and maintenance of fully functional electron transport.
A number of functions are served by mitochondrial fission. For example, mitochondrial fission is particularly active when cells divide (particularly during the G2 and M phases of the cell cycle; see Chapter 19). As a consequence, the multiple discrete mitochondria generated by fission are readily distributed evenly into the daughter cells. Mitochondria can be transported by motor proteins along cytoskeletal filaments, including microtubules and microfilaments (see Chapter 17), to establish their proper intracellular distribution. In addition, fission provides a powerful mechanism of quality control by culling defective segments of mitochondria from the interconnected, healthy mitochondrial network. Should a portion of a large mitochondrial network become damaged or dysfunctional—for example, by the generation of high levels of reactive oxygen species (discussed later) or by mutations in the mitochondrial DNA—fission can separate the compromised segments from healthy segments.
Cells have the capacity to recognize damaged or dysfunctional segments of the mitochondrial network and, after they detach from the network, can surround them with a membrane and then deliver them to lysosomes for degradation. This destruction of mitochondria, which is called mitophagy (“eating mitochondria”), is a subset of the general process called autophagy (“self eating”) by which cells engulf in membranes and degrade organelles and portions of the cytosol (see Chapter 14). Strikingly, hereditary early-onset Parkinson’s disease is caused by mutations in two genes encoding proteins that can mediate mitophagy. The proteins are PINK1 (a kinase) and Parkin (an E3 ubiquitin ligase that covalently links the small protein ubiquitin to nearby proteins to target them for destruction by proteasomes; see Chapter 3). In healthy mitochondria, PINK1 is imported into the mitochondrial matrix. When the mitochondrion is damaged or dysfunctional, PINK1 cannot enter the matrix and remains at the outer mitochondrial membrane, where it recruits cytosolic Parkin to the damaged segment. The Parkin is activated and polyubiquitinylates outer-membrane proteins, thus targeting them for degradation and inducing mitophagy. When the environment of a cell has inadequate oxygen (hypoxia), certain enzymes modify proteins on the outer mitochondrial membrane, inducing mitophagy. The influences of PINK1 and Parkin on mitochondrial homeostasis, which may include mitophagy-independent as well as mitophagy-dependent pathways, were first identified in genetic studies of the fly Drosophila melanogaster.
Mitochondrial dynamism has additional features that are worth mentioning. Mitochondrial structure and function can change in response to the metabolic state of the cells. For example, the isolation of rat liver cells (hepatocytes) from the body and their transfer into cell culture stresses the cells and results in their depolarization (loss of some of their epithelial cell properties; see Chapter 20). These cells exhibit fragmentation of their mitochondria (Figure 12-12d, left) accompanied by low generation of ATP via oxidative phosphorylation. As the cells adjust to growth in cell culture, they become polarized (have a morphology and biochemistry more like that of hepatocytes in a liver), increase mitochondrial generation of ATP via oxidative phosphorylation, and exhibit an extensive network of fused mitochondria (Figure 12-12d, right). Remarkably, recent studies have suggested that mtDNA, and indeed intact mitochondria, can be transferred from one cell to another via membrane tubules called tunneling nanotubes, which are described in Chapter 20.