11-2 Motor System Organization
Although we humans tend to rely primarily on our hands for manipulating objects, we can still learn to handle things with other body parts, such as the mouth or a foot, if we have to. Some people without arms become proficient at using a foot for writing or painting or even for driving. What properties of the motor system allow such versatility in carrying out such skilled movements? In this section we examine first the organization of the motor cortex, then the descending pathways from the motor cortex to the brainstem and spinal cord, and finally the motor neurons that in turn connect with the body’s muscles.
Motor Cortex
Section 4-1 describes the milestones that led to understanding how the nervous system uses electrical charge to convey information.
In 1870, two Prussian physicians, Gustav Fritsch and Eduard Hitzig, discovered that they could electrically stimulate the neocortex of an anesthetized dog to produce movements of the mouth, limbs, and paws on the opposite side of the dog’s body. They provided the first direct evidence that the neocortex controls movement. Later researchers confirmed the finding by experimenting with a variety of animals as subjects, including rats, monkeys, and apes.
Figure 10-19 shows Penfield using brain stimulation to map the cortex.
Based on this research background, beginning in the 1930s Wilder Penfield (Penfield & Boldrey, 1958) used electrical stimulation to map the cortices of conscious human patients who were about to undergo neurosurgery. Penfield’s aim was to use the results to assist in surgery. He and his colleagues confirmed that movements in humans are triggered mainly in response to stimulation of the premotor and primary motor cortices.
Mapping the Motor Cortex
Penfield summarized his results by drawing cartoons of body parts to represent the areas of the motor cortex that produce movement in those parts. The result was a homunculus (pl. homunculi; Latin for little person) that could be spread out across the primary motor cortex, as illustrated in Figure 11-6. Because the body is symmetrical, an equivalent motor homunculus is discernible in the primary motor cortex of each hemisphere, and each motor cortex mainly controls movement in the opposite side of the body. Penfield also identified another, smaller motor homunculus in the dorsal premotor area of each frontal lobe, a region sometimes referred to as the supplementary motor cortex.
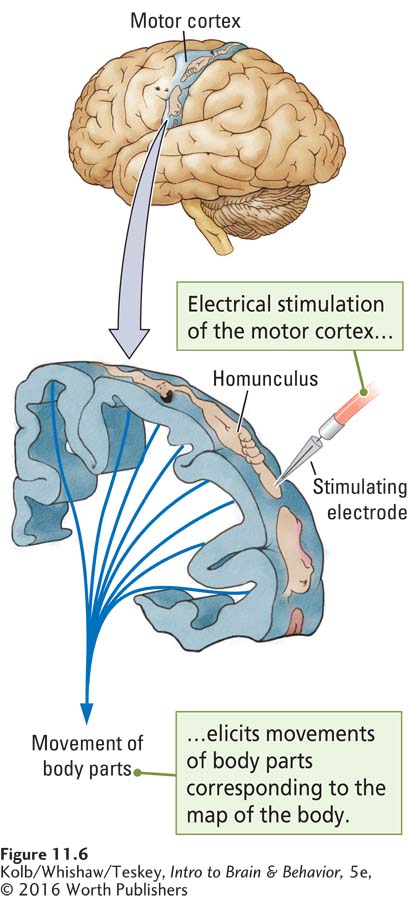
The striking feature of the homunculus shown in Figure 11-7 is the disproportionate relative sizes of its body parts compared with the relative sizes of actual parts of the human body. The homunculus has huge hands with an especially large thumb. Its lips and tongue are also prominent. By contrast, the trunk, arms, and legs—
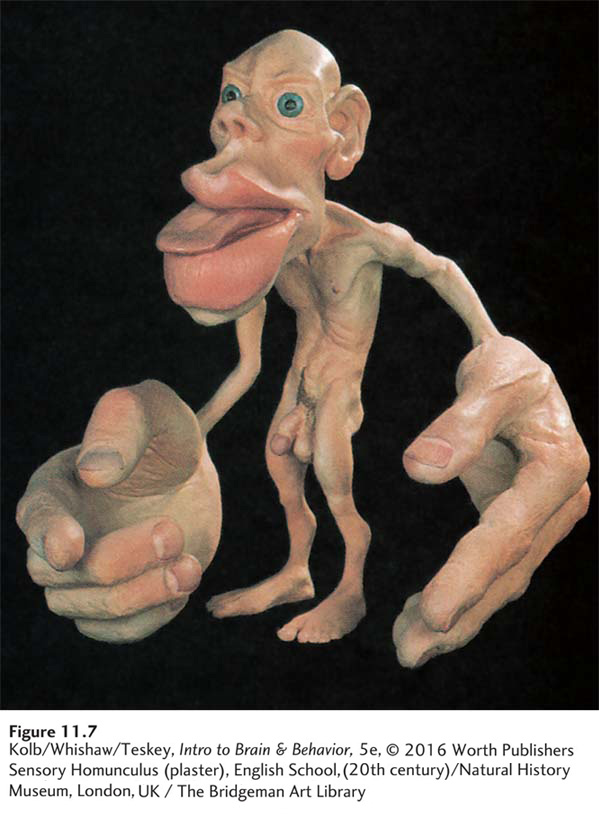
These distortions illustrate that extensive areas of M1 allow precise regulation of the hands, fingers, lips, and tongue (see Figure 11-6). Body areas over which we have relatively little motor control have a much smaller representation in the motor cortex.
Another curious feature of the homunculus as laid out across the motor cortex is that the body parts are discontinuous—
Modeling Movement
The motor homunculus shows at a glance that relatively large areas of the brain control the body parts we use to make the most skilled movements—
An early idea was that each part of the homunculus controls muscles in that part of the body. Information from other cortical regions could be sent to the motor homunculus, and neurons in the appropriate part of the homunculus could then activate body muscles required for producing the movement. If you wanted to pick up a coin, for example, messages from the M1 finger area would instruct the fingers. More recent experiments suggest that the motor cortex represents not muscles but rather a repertoire of fundamental movement categories (Grazaino, 2006).
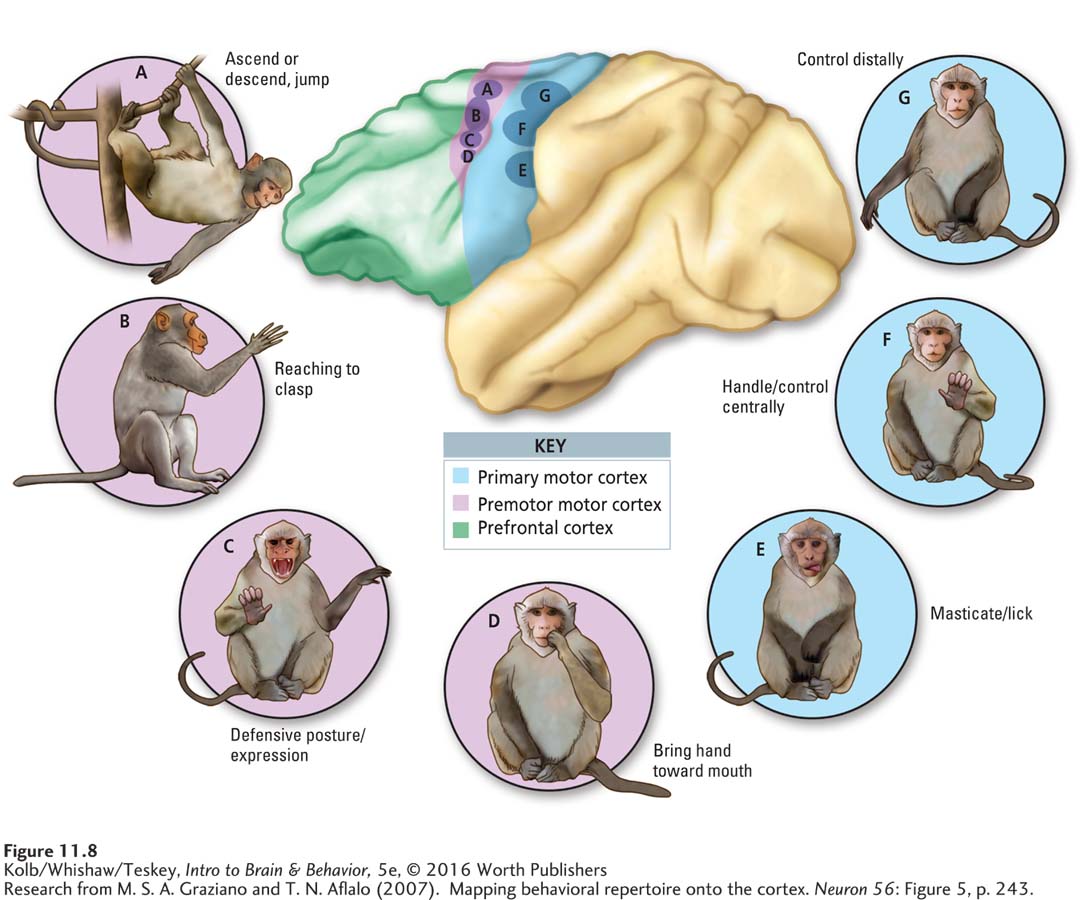
The drawings in Figure 11-8 illustrate several movement categories elicited in monkeys by electrical stimulation. They include (A) ascend, descend, or jump, (B) reach to clasp, (C) defensive posture or expression, (D) hand toward mouth, (E) masticate or lick, (F) control centrally, and (G) control distally. Whole-
Each observed movement has the same end regardless of the starting location of a monkey’s limb or its other ongoing behavior. Electrical stimulation that results in the hand coming to the mouth always recruits the hand. If a weight is attached to the monkey’s arm, the evoked movement compensates for the added load.
But categorized movements are inflexible: when an obstacle is placed between the hand and the mouth, the hand hits the obstacle. If stimulation continues after the hand has reached the mouth, the hand remains there for the duration of the stimulation. Further, broad movement categories—
MRI studies on human subjects suggest that the human motor cortex, like the monkey motor cortex, is organized in terms of functional movement categories (Meier et al., 2008). The motor cortex maps appear to represent basic types of movements that learning and practice can modify. In other words, the motor cortex encodes not muscle twitches but a lexicon, or dictionary, of movements. As with words and sentences, these few movements used in different combinations produce all the movements you are capable of, even in activities as complex as playing basketball.
Motor Cortex and Skilled Movement
In a study designed to investigate how neurons in the motor cortex control movement, Edward Evarts (1968) used the simple procedure illustrated in Experiment 11-2. He trained a monkey to flex its wrist to move a bar. Different weights could be attached to the bar. An electrode implanted in the wrist region of the monkey’s motor cortex recorded the activity of neurons there.
EXPERIMENT
Question: How does the motor cortex take part in the control of movement?
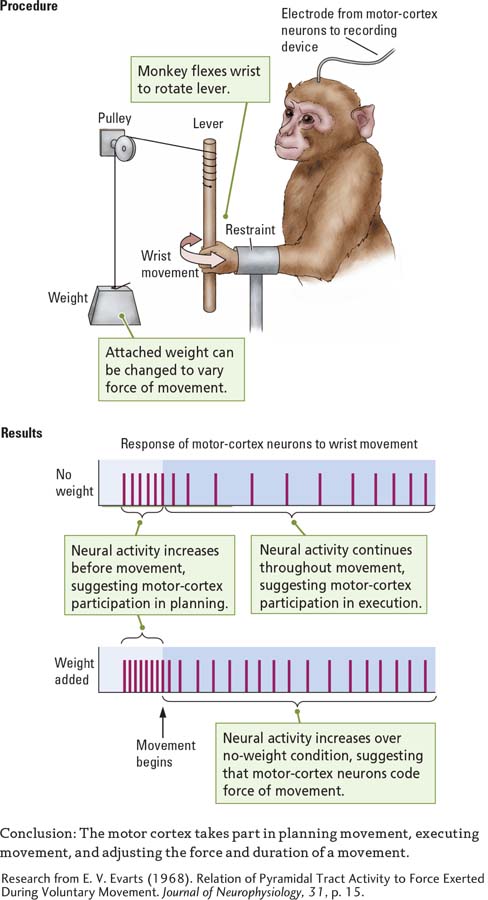
Evarts discovered that the neurons began to discharge even before the monkey began the movement, as shown in the Results section of Experiment 11-2. Thus, they take part in planning the movement as well as initiating it. The neurons continued to discharge as the wrist moved, confirming that they play a role in producing the movement. Finally, the neurons discharged at a higher rate when the bar was loaded with a weight. This finding shows that motor cortex neurons increase a movement’s force by increasing their firing rate and its duration, as stated in the experiment’s Conclusion.
Evarts’s findings also reveal that the motor cortex has a role in specifying the end point of a movement. Motor cortex neurons in the wrist might discharge when the monkey flexed its wrist inward but not when the wrist was extended back to its starting position. These on–
The activity of motor neurons in freely moving monkeys suggests that the neurons, depending upon their location, move the body or parts of the body to specific positions (Aflalo & Graziano, 2007). For example, if some neurons are very active when a hand is moved to a specific location, they are progressively less active when the hand is moved away from that position. Thus, the neurons in the motor cortex regions illustrated in Figure 11-8 are responsible for configuring movements that achieve the same end point produced when those same neurons are electrically stimulated (Aflalo & Graziano, 2007).
Nevertheless, studies using human participants reveal situations in which motor cortex neurons can be active even when no overt movement occurs (Schieber, 2011). These situations include planning a movement, withholding a movement on instruction, and mental imagery. Such subthreshold activity in motor cortex neurons may underlie our ability to imagine movements without actually producing them. Subthreshold activity may also allow motor cortex neurons to control brain–
Plasticity in the Motor Cortex
Section 14-4 explores how motor maps change in response to learning.
An intimate relationship connects the activity of M1 neurons with movement of the body. The studies just described show that flexibility is part of the relationship. Flexibility underlies another property of the motor cortex: its plasticity, which contributes both to motor learning and to recovery after the motor cortex is damaged, as the following example explains.
Focus 2-3 describes motor disruptions that stroke causes; Section 16-3 reviews stroke treatments.
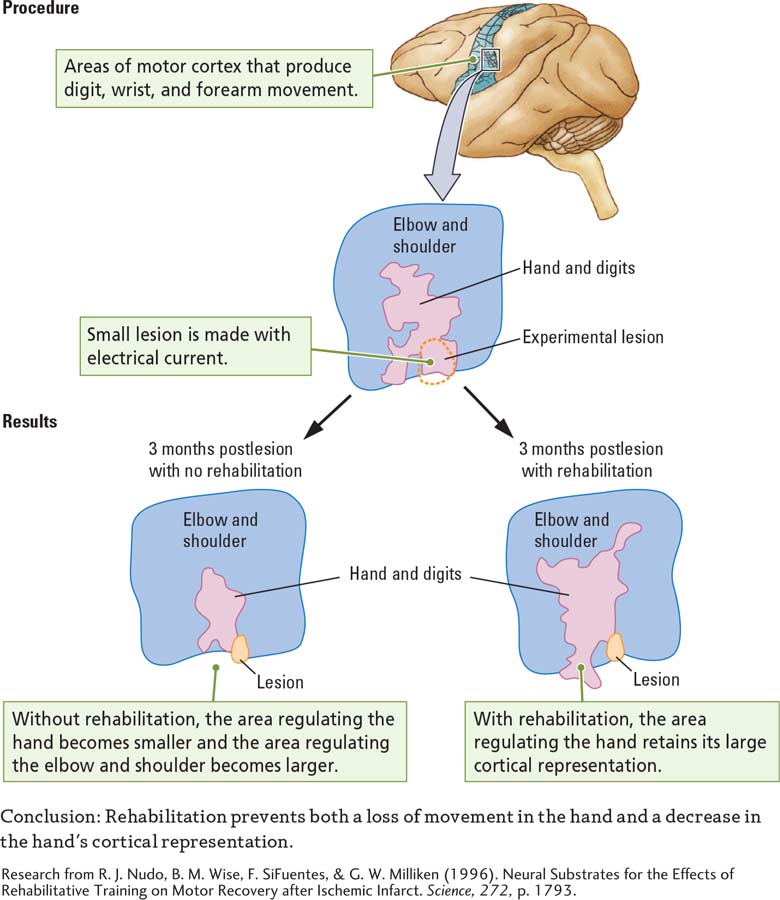
A study by Randy Nudo and his coworkers (1996), summarized in the Procedure section of Experiment 11-3, illustrates change due to cortical damage. These researchers mapped the motor cortices of monkeys to identify the hand and digit areas. They then surgically removed a small part of the cortex that represents the digit area. After undergoing this electrolytic lesion, the monkeys used the affected hand much less, relying mainly on the good hand.
EXPERIMENT
Question: What is the effect of rehabilitation on the cortical representation of the forelimb after brain damage?
Section 7-1 describes ablation techniques used by neuroscience researchers to manipulate the brain.
When the researchers stimulated the cortex of the monkeys 3 months later, the animals were unable to produce many lower-
The experimenters wondered whether the change could have been prevented had they forced the monkeys to use the affected arm. To find out, they used the same procedure on other monkeys, except that during the postsurgery period they forced the animals to rely on the bad arm by binding the good arm in a sling.
When the experimenters reexamined these monkeys’ motor maps 3 months later, they found that the hand and digit area retained its larger size. Even though no neural activity occurred in the spot with the lesion, the monkeys had gained some function in the digits that used to be connected to the damaged spot. Apparently, the remaining cortical digit area was now controlling the movement of these fingers.
Most likely, plasticity is promoted in the formation of new connections and the strengthening of existing connections among different parts of the motor homunculus. Humans who have had a stroke to the motor cortex also display plasticity-
As Nudo’s monkey experiment illustrates, one way to enhance recovery is to restrain the good limb. Constraint-
Corticospinal Tracts
The main efferent pathways from the motor cortex to the brainstem to the spinal cord are the corticospinal tracts. The axons from these tracts originate mainly in motor cortex layer V pyramidal cells but also extend from the premotor cortex and the sensory cortex (see Layering in the Neocortex on page 359). The axons descend into the brainstem, sending collaterals to numerous brainstem nuclei, and eventually emerge on the brainstem’s ventral surface, where they form a large bump on each side. These bumps, or pyramids, give the corticospinal tracts their alternative name, the pyramidal tracts.
At this point, some axons descending from the left hemisphere cross over to the right side of the brainstem. Likewise, some axons descending from the right hemisphere cross over to the left side of the brainstem. The remaining axons stay on their original side. This division produces two corticospinal tracts, one crossed and the other uncrossed, entering each side of the spinal cord. Figure 11-9 illustrates the division of tracts originating in the left-
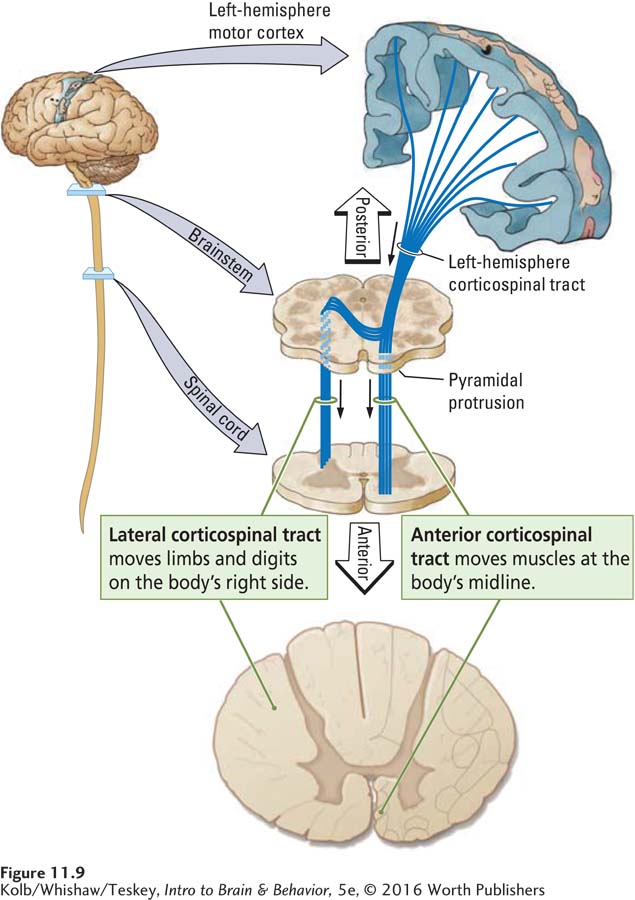
The cross section of a spinal cord in Figure 11-10 shows the location of the two tracts, on the left and right sides. The fibers that cross to the opposite side of the brainstem descend the spinal cord in a lateral (side) position to form the lateral corticospinal tract. The fibers that remain on their original side continue from the brainstem down the spinal cord in an anterior (front) position, to form the anterior corticospinal tract.
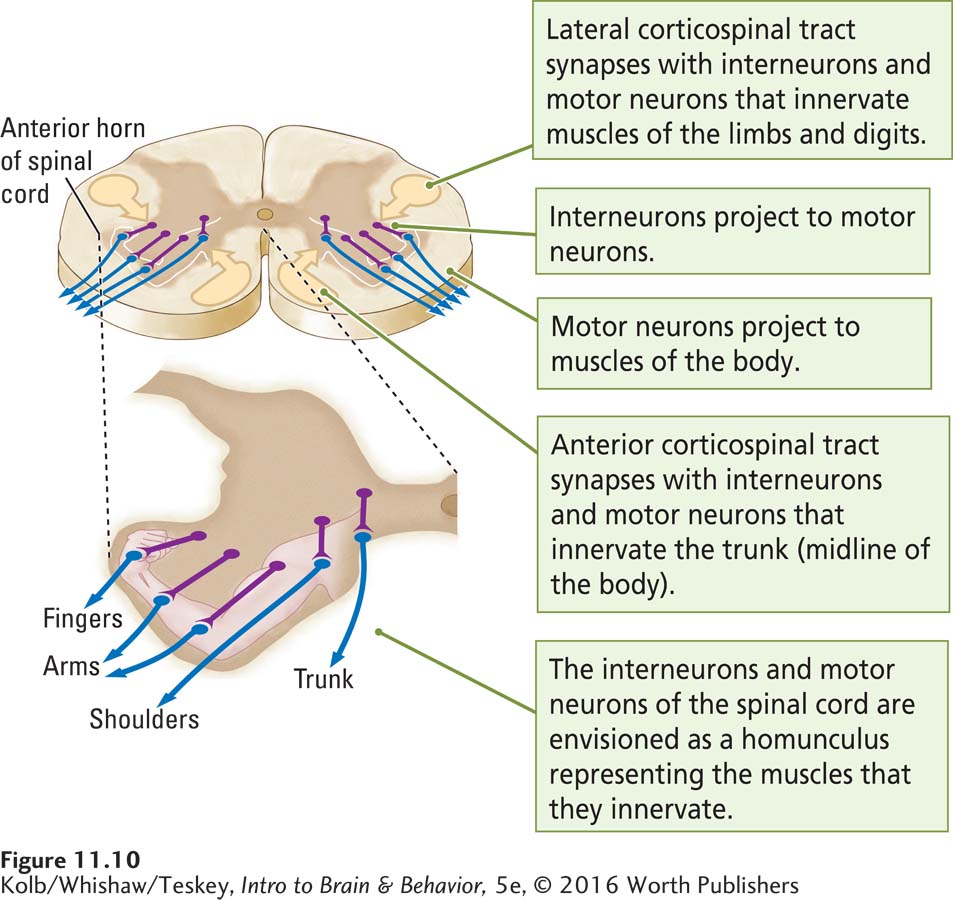
Retracing the pathway, the corticospinal tracts originate in the neocortex and terminate in the spinal cord. Within the spinal cord, corticospinal fibers make synaptic connections with both interneurons and motor neurons, but the motor neurons carry all nervous system commands out to the muscles.
Motor Neurons
Spinal cord motor neurons are located in the anterior part of the spinal cord, the anterior horns, which jut out from the anterior part of the spinal cord. The anterior horns contain two kinds of neurons. Interneurons lie just medial to the motor neurons and project onto them. The motor neurons send their axons to the body muscles. The fibers from the corticospinal tracts make synaptic connections with both the interneurons and the motor neurons, but the motor neurons carry all nervous system commands to the muscles.
Figure 11-10 shows that a homunculus of the body is represented again in the spinal cord. The more lateral motor neurons project to muscles that control the fingers and hands, whereas intermediate motor neurons project to muscles that control the shoulders and arms. The most medial motor neurons project to muscles that control the body’s trunk. Axons of the lateral corticospinal tract connect mainly with the lateral interneurons and motor neurons, and axons of the anterior corticospinal tract connect mainly to the medial interneurons and motor neurons.
To visualize how the cortical regions responsible for different movements relate to the motor neuron homunculus in the spinal cord, look again at Figure 11-9. Place your finger on the index finger region of the motor homunculus on the left side of the brain. If you trace the axons of the cortical neurons downward, your route takes you through the brainstem, across its midline, and down the right lateral corticospinal tract.
If you are right-
The journey ends at the interneurons and motor neurons in the most lateral region of the spinal cord’s right anterior horn—
If you repeat the procedure by tracing the pathway from the trunk area of the motor homunculus, near the top on the left side of the brain, you follow the same route through the upper part of the brainstem. You do not cross over to the opposite side, however. Instead, you descend into the spinal cord on the left side, the same (ipsilateral) side of the nervous system on which you began, eventually ending up in the most medial interneurons and motor neurons of the left side’s anterior horn. (At this point, some of these axons also cross over to the other side of the spinal cord.) Thus, if you follow these motor neuron axons, you end up at their synapses with the muscles that move the trunk on both sides of the body.
This visualization can help you remember the routes taken by motor system axons. The limb regions of the motor homunculus contribute most of their fibers to the lateral corticospinal tract, the fibers that cross over to the opposite side of the spinal cord. They activate motor circuits that move the arm, hand, leg, and foot on the opposite side of the body. In contrast, the trunk regions of the motor homunculus contribute their fibers to the anterior corticospinal tract. Only a few of these fibers cross, close to their termination in the spinal cord; most control the trunk and limbs on the same side of the body.
Remember that the motor cortex is organized in terms of functional movement categories, such as reaching or climbing (see Figure 11-8). A similar template in the spinal cord ensures that instructions from the motor cortex are reproduced faithfully. Presumably, lateral interneurons produce acts of reaching or bringing the hand to the mouth, and the medial interneurons and motor neurons produce whole-
In addition to the corticospinal pathways, about 24 other pathways from the brainstem to the spinal cord carry instructions, such as information related to posture and balance (see Section 11-4), and they control the enteric nervous system as well as portions of the sympathetic division of the ANS. Remember that for all these functions, the motor neurons are the final common path.
Control of Muscles
Spinal cord motor neurons synapse on the muscles that control body movements. For example, the biceps and triceps of the upper arm control movement of the lower arm. Limb muscles are arranged in pairs, as shown in Figure 11-11. One member of a pair, the extensor, moves (extends) the limb away from the trunk. The other member of the pair, the flexor, moves (flexes) the limb in toward the trunk. Experiment 11-2 on page 368 demonstrates the on–
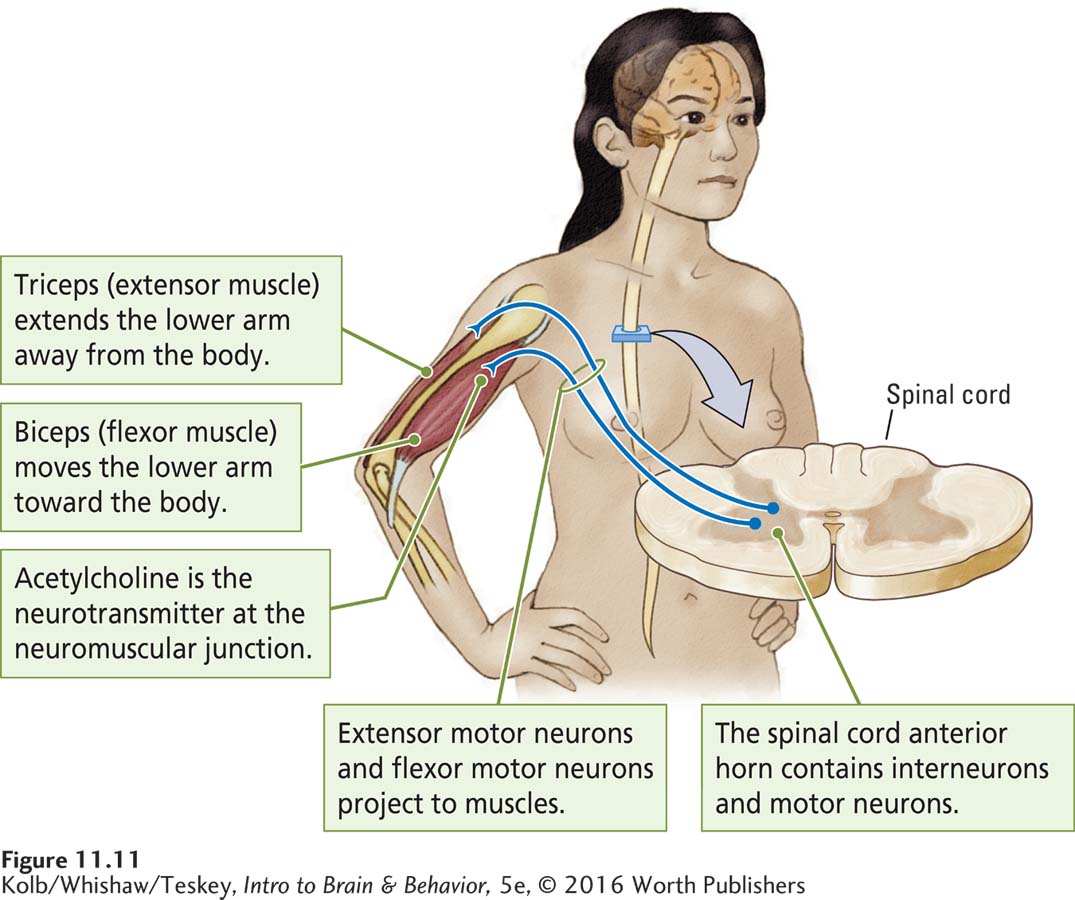
Figure 4-26 illustrates ACh action at a motor neuron–
Connections between spinal cord interneurons and motor neurons ensure that the muscles work together so that when one muscle contracts, the other relaxes. Thus, the spinal cord interneurons and motor neurons not only relay instructions from the brain but also, through their connections, cooperatively organize the movement of many muscles. As you know, the neurotransmitter at the motor neuron–
11-2 REVIEW
Motor System Organization
Before you continue, check your understanding.
Question 1
The ____________ organization of the motor cortex is represented by a ____________, in which parts of the body that are capable of the most skilled movements (especially the mouth, fingers, and thumbs) are regulated by ____________ cortical regions.
Question 2
Change can take place in the cortical ____________ to aid in recovery of function after motor cortex injury.
Question 3
Instructions regarding movement travel out from the motor cortex through the ____________ tracts to terminate on interneurons that project to motor neurons in the anterior horn of the spinal cord. Many corticospinal-
Question 4
The anterior corticospinal tracts carry instructions for ____________ movements, whereas the lateral corticospinal tracts carry instructions for ____________ and ____________ movements.
Question 5
Motor neuron axons in the spinal cord carry instructions to ____________ that are arranged in pairs. One ____________ a limb; the other ____________ the limb.
Question 6
What does the plan of movements in the motor cortex as revealed by electrical stimulation tell us about the brain’s representation of movement?
Answers appear in the Self Test section of the book.