5-4 Adaptive Role of Synapses in Learning and Memory
Among our most cherished abilities are learning and remembering. Neuroplasticity is a requirement for learning and memory and a characteristic of the mammalian brain. In fact, it is a trait of the nervous systems of all animals, even the simplest worms. Larger brains with more connections are more plastic, however, and thus likely to show more adaptability in neural organization.
Experiment 2-1 demonstrates observational learning in the octopus and the ubiquity of neuroplasticity.
Greater adaptability happens because experience can alter the synapse. Not only are synapses versatile in structure and function, they are plastic: they can change. The synapse, therefore, is the site for the neural basis of learning, a relatively permanent change in behavior that results from experience.
Hebb’s “cell-
Donald O. Hebb (1949) was not the first to suggest that learning is mediated by structural changes in synapses. But the change that he envisioned in his book The Organization of Behavior was novel 65 years ago. Hebb theorized, “When an axon of cell A is near enough to excite a cell B and repeatedly or persistently takes part in firing it, some growth process or metabolic change takes place in one or both cells such that A’s efficiency, as one of the cells firing B, is increased” (Hebb, 1949, p. 62). Simply put, cells that fire together wire together. A synapse that physically adapts in this way is called a Hebb synapse today.

Eric Kandel was awarded a Nobel Prize in 2000 for his descriptions of the synaptic basis of learning in a way that Hebb envisaged: learning in which the conjoint activity of nerve cells serves to link them. Kandel’s subject, the marine slug Aplysia californica, is an ideal subject for learning experiments. Slightly larger than a softball and lacking a shell, Aplysia has roughly 20,000 neurons. Some are quite accessible to researchers, who can isolate and study circuits having few synapses.
When threatened, Aplysia defensively withdraws its more vulnerable body parts—
Section 14-4 investigates the neural bases of brain plasticity in conscious learning and in memory.
We illustrate the role of synapses in two kinds of learning that Kandel has studied: habituation and sensitization. For humans, both are called unconscious because they do not depend on a person’s knowing precisely when and how they occur.
Habituation Response
In habituation, the response to a stimulus weakens with repeated stimulus presentations. If you are accustomed to living in the country, then move to a city, you might at first find the sounds of traffic and people extremely loud and annoying. With time, however, you stop noticing most of the noise most of the time. You have habituated to it.
Habituation develops with all our senses. When you first put on a shoe, you feel it on your foot, but very soon it is as if the shoe were not there. You have not become insensitive to sensations, however. When people talk to you, you still hear them; when someone steps on your foot, you still feel the pressure. Your brain simply has habituated to the customary background sensation of a shoe on your foot.
Aplysia habituates to waves in the shallow tidal zone where it lives. These slugs are constantly buffeted by the flow of waves against their body, and they learn that waves are just the background noise of daily life. They do not flinch and withdraw every time a wave passes over them. They habituate to this stimulus.
A sea slug that is habituated to waves remains sensitive to other touch sensations. Prodded with a novel object, it responds by withdrawing its siphon and gill. The animal’s reaction to repeated presentations of the same novel stimulus forms the basis for Experiment 5-2, studying its habituation response.
Neural Basis of Habituation
The Procedure section of Experiment 5-2 shows the setup for studying what happens to the withdrawal response of Aplysia’s gill after repeated stimulation. A gentle jet of water is sprayed on the siphon while gill movement is recorded. If the water jet is presented to Aplysia’s siphon as many as 10 times, the gill withdrawal response is weaker some minutes later, when the animal is again tested. The decrement in the strength of the withdrawal is habituation, which can last as long as 30 minutes.
EXPERIMENT
Question: What happens to the gill response after repeated stimulation?
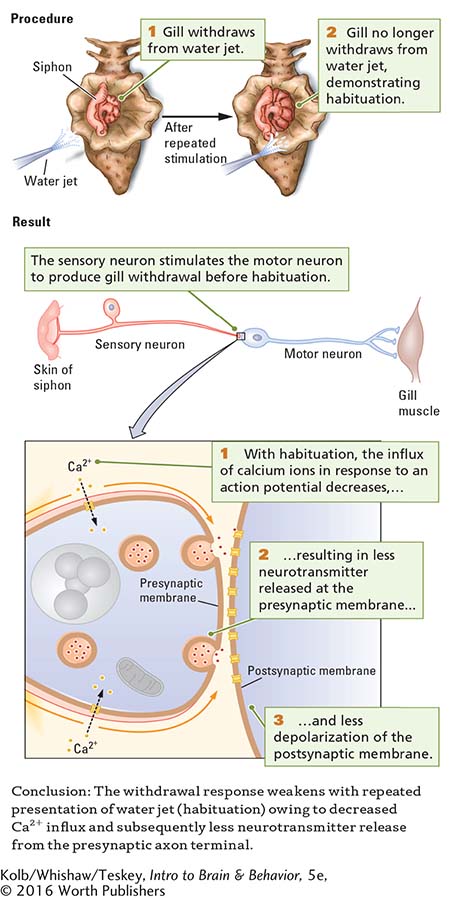
The Results section of Experiment 5-2 starts by showing a simple representation of the pathway that mediates Aplysia’s gill withdrawal response. For purposes of illustration, only one sensory neuron, one motor neuron, and one synapse are shown; in actuality, about 300 neurons may take part in this response. The water jet stimulates the sensory neuron, which in turn stimulates the motor neuron responsible for the gill withdrawal. But exactly where do the changes associated with habituation take place? In the sensory neuron? In the motor neuron? In the synapse between the two?
Habituation does not result from an inability of either the sensory or the motor neuron to produce action potentials. In response to direct electrical stimulation, both the sensory neuron and the motor neuron retain the ability to generate action potentials even after habituation. Electrical recordings from the motor neuron show that as habituation develops, the excitatory postsynaptic potentials (EPSPs) in the motor neuron become smaller.
The most likely way in which these EPSPs decrease in size is that the motor neuron is receiving less neurotransmitter from the sensory neuron across the synapse. And if less neurotransmitter is being received, then the changes accompanying habituation must be taking place in the presynaptic axon terminal of the sensory neuron.
Reduced Sensitivity of Calcium Channels Underlies Habituation
Kandel and his coworkers measured neurotransmitter output from a sensory neuron and verified that less neurotransmitter is in fact released from a habituated neuron than from a nonhabituated one. Recall from Figure 5-5 that neurotransmitter release in response to an action potential requires an influx of calcium ions across the presynaptic membrane. As habituation takes place, that Ca2+ influx decreases in response to the voltage changes associated with an action potential. Presumably, with repeated use, voltage-
The neural basis of habituation lies in the change in presynaptic calcium channels. Its mechanism, which is summarized close up in the Results section of Experiment 5-2, is a reduced sensitivity of calcium channels and a consequent decrease in neurotransmitter release. Thus, habituation can be linked to a specific molecular change, as summarized in the experiment’s Conclusion.
Sensitization Response
A sprinter crouched in her starting blocks is often hyperresponsive to the starter’s gun: its firing triggers in her a rapid reaction. The stressful, competitive context of the race helps to sensitize the sprinter to this sound. Sensitization, an enhanced response to some stimulus, is the opposite of habituation. The organism becomes hyperresponsive to a stimulus rather than accustomed to it.
Sensitization occurs within a context. Sudden, novel stimulation heightens our general awareness and often results in larger-
Stress can foster and prolong PTSD effects. See Sections 6-5 and 12-4. Section 16-4 covers treatment strategies.
The same thing happens to Aplysia. Sudden, novel stimuli can heighten a slug’s responsiveness to familiar stimulation. When attacked by a predator, for example, the slug displays heightened responses to many other stimuli in its environment. In the laboratory, a small electric shock to Aplysia’s tail mimics a predatory attack and effects sensitization, as illustrated in the Procedure section of Experiment 5-3. A single electric shock to the slug’s tail enhances its gill withdrawal response for a period that lasts for minutes to hours.
EXPERIMENT
Question: What happens to the gill response in sensitization?
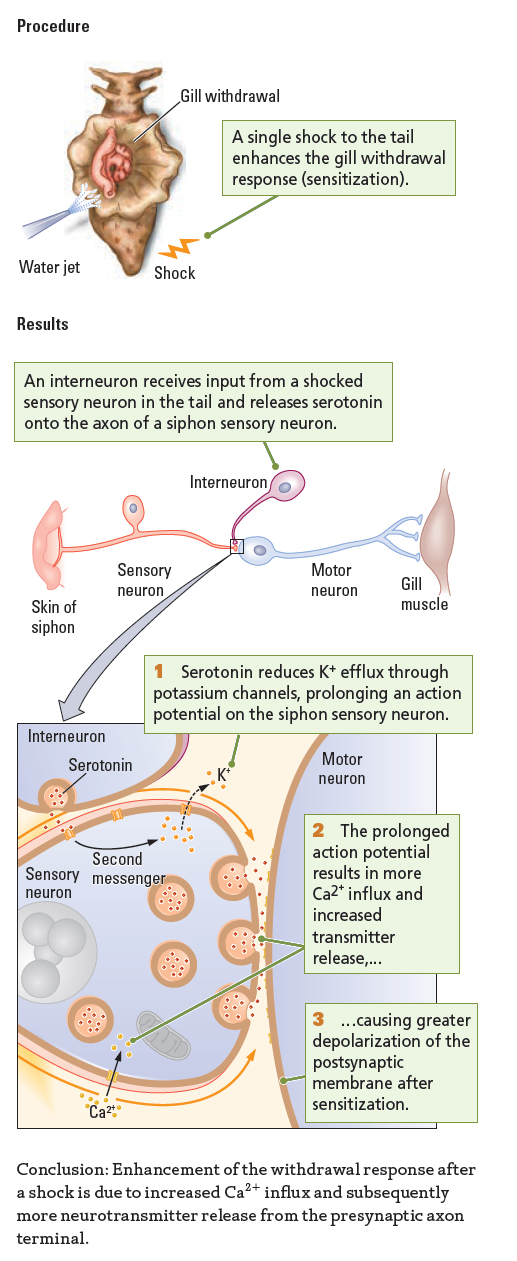
Neural Basis of Sensitization
The neural circuits participating in sensitization differ from those that take part in a habituation response. The Results section of Experiment 5-3 shows the sensory and motor neurons that produce the gill withdrawal response and adds an interneuron that is responsible for sensitization.
An interneuron that receives input from a sensory neuron in Aplysia’s tail (and so carries information about the shock) makes an axoaxonic synapse with a sensory neuron in the siphon. The interneuron’s axon terminal contains serotonin. Consequently, in response to a tail shock, the tail sensory neuron activates the interneuron, which in turn releases 5-
At the molecular level, shown close up in Experiment 5-3 Results, the serotonin released from the interneuron binds to a metabotropic serotonin receptor on the siphon’s sensory neuron axon. This binding activates second messengers in the sensory neuron. Specifically, the serotonin receptor is coupled through its G protein to the enzyme adenyl cyclase. This enzyme increases the concentration of second messenger cyclic adenosine monophosphate (cAMP) in the presynaptic membrane of the siphon’s sensory neuron.
Through several chemical reactions, cAMP attaches a phosphate molecule (PO4) to potassium channels, rendering them less responsive. The close-
Less-
The longer-
This increased neurotransmitter release produces greater activation of the motor neuron and thus a larger-
Sensitization, then, is the opposite of habituation at the molecular level as well as at the behavioral level. In sensitization, more Ca2+ influx results in more transmitter being released, whereas in habituation, less Ca2+ influx results in less neurotransmitter being released. The structural basis of cellular memory in these two forms of learning is different, however. In sensitization, the change takes place in potassium channels, whereas in habituation, the change takes place in calcium channels.
Learning as a Change in Synapse Number
Neural changes associated with learning must last long enough to account for a relatively permanent change in an organism’s behavior. The changes at synapses described in the preceding sections develop quite quickly, but they do not last indefinitely, as memories often do. How then, can synapses be responsible for the long-
Repeated stimulation produces habituation and sensitization that can persist for months. Brief training produces short-
Researchers working with Eric Kandel (Bailey et al., 2015) found that the number and size of sensory synapses change in well-
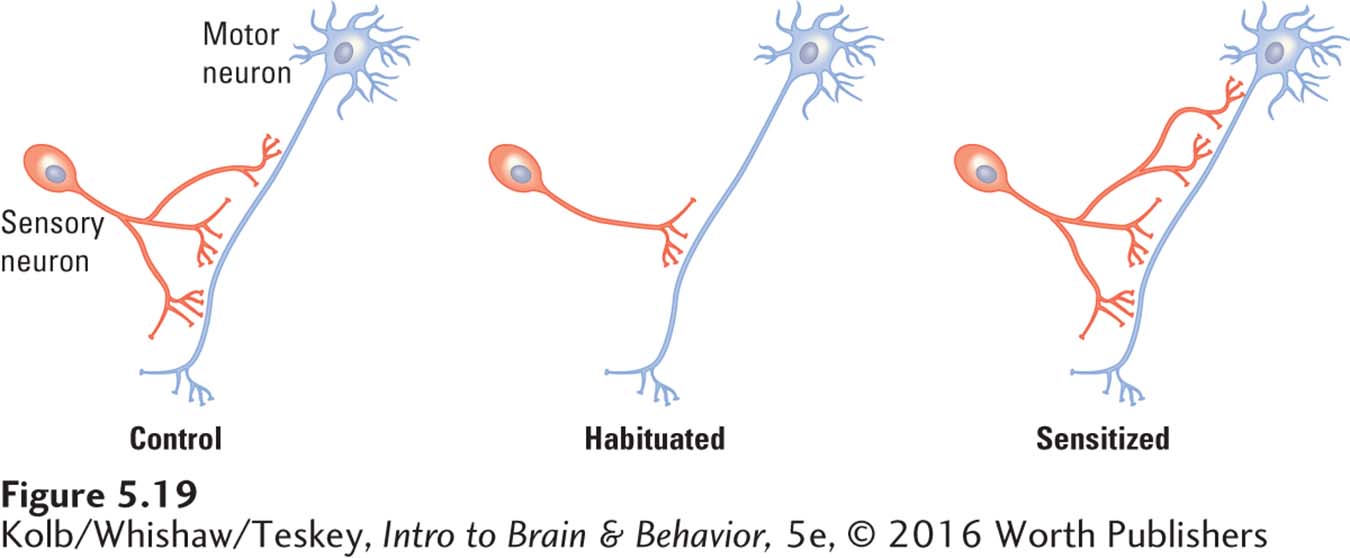
A mechanism through which these processes can take place begins with calcium ions that mobilize second messengers to send instructions to nuclear DNA. The transcription and translation of nuclear DNA in turn initiate structural changes at synapses, including the formation of new synapses and new dendritic spines. Research Focus 5-5, Dendritic Spines: Small but Mighty, summarizes experimental evidence about structural changes in dendritic spines.
The second messenger cAMP plays an important role in carrying instructions regarding these structural changes to nuclear DNA. The evidence for cAMP’s involvement comes from studies of the fruit fly Drosophila. Two genetic mutations in the fruit fly can produce similar learning deficiencies. Both render the second messenger cAMP inoperative, but in opposite ways. One mutation, called dunce, lacks the enzymes necessary to degrade cAMP, so the fruit fly has abnormally high cAMP levels. The other mutation, called rutabaga, reduces levels of cAMP below the normal range for Drosophila neurons.
Significantly, fruit flies with either mutation are impaired in acquiring habituated and sensitized responses because their levels of cAMP cannot be regulated. New synapses seem to be required for learning to take place, and the second messenger cAMP seems to carry instructions to form them. Figure 5-20 summarizes these research findings.

5-5
Dendritic Spines: Small but Mighty
Dendritic spines, which protrude from the dendrite’s shaft, measure about 1 to 3 micrometers (μm, one-
Dendritic spines originate in filopodia (from the Latin file, for thread, and the Greek podium, for foot) that bud out of neurons, especially at dendrites. Microscopic observation of dendrites shows that filopodia are constantly emerging and retracting over times on the order of seconds.
This budding of filopodia is much more pronounced in developing neurons and in the developing brain (see Figure 8-13). Because filopodia can grow into dendritic spines, their budding suggests that they are searching for contacts from axon terminals to form synapses. When contact is made, some new synapses may have only a short life; others will endure.
A permanent dendritic spine tends to have a large, mushroom-
Dendritic spines mediate learning that lasts, including habituation and sensitization. To mediate learning, each spine must be able to act independently, undergoing changes that its neighbors do not undergo.
Examination of dendritic spines in the nervous system shows that some are simple and others complex. The cellular mechanisms that allow synapses to appear on spines and to change shape include microfilaments linked to the membrane receptors, protein transport from the cell body, and the incorporation of nutrients from the extracellular space.
The variety suggests that all this activity changes the appearance of both presynaptic and postsynaptic structures. The illustration summarizes synaptic structures that can be measured and related to learning and behavior and to structural changes that may subserve learning.
Dendritic spines provide the structural basis for our behavior, our individual skills, and our memories (Bosch & Hayashi, 2012). Impairments in forming spines characterize some kinds of mental disability, and the loss of spines is associated with the dementia of Alzheimer disease.
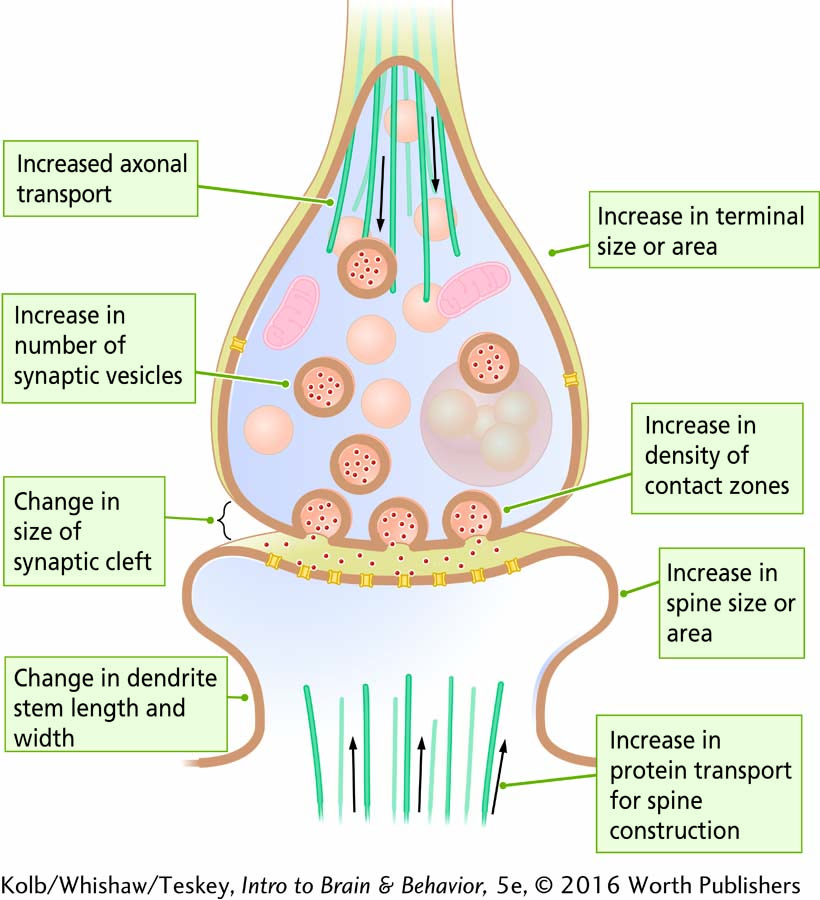
More lasting habituation and sensitization are mediated by relatively permanent changes in neuronal structure—
5-4 REVIEW
Adaptive Role of Synapses in Learning and Memory
Before you continue, check your understanding.
Question 1
Experience alters the _______, the site of the neural basis of _______, a relatively permanent change in behavior that results from experience.
Question 2
Aplysia’s synaptic function mediates two basic forms of learning: _______and _______.
Question 3
Changes that accompany habituation take place within the _______ of the _______ neuron, mediated by _______ channels that grow _______ sensitive with use.
Question 4
The sensitization response is amplified by _______ that release serotonin onto the presynaptic membrane of the sensory neuron, changing the sensitivity of presynaptic _______ channels and increasing the influx of _______.
Question 5
One characteristic of _______, defined as physiological arousal related to recurring memories and dreams surrounding a traumatic event that persist for months or years after the event, is a heightened response to stimuli. This suggests that the disorder is in part related to _______.
Question 6
Describe the benefits and/or drawbacks of permanent habituation and sensitization.
Answers appear in the Self Test section of the book.