Membrane Channels and Pumps
CHAPTER
13
367
OUTLINE
The lipid bilayer of biological membranes is intrinsically impermeable to ions and polar molecules, yet these species must be able to cross these membranes for normal cell function. Permeability is conferred by three classes of membrane proteins: pumps, carriers, and channels. Pumps use a source of free energy such as ATP hydrolysis or light absorption to drive the thermodynamically uphill transport of ions or molecules. Pump action is an example of active transport. Carriers mediate the transport of ions and small molecules across the membrane without consumption of ATP. Channels provide a membrane pore through which ions can flow very rapidly in a thermodynamically downhill direction. The action of channels illustrates passive transport, or facilitated diffusion.
Pumps are energy transducers in that they convert one form of free energy into another. Two types of ATP-
368
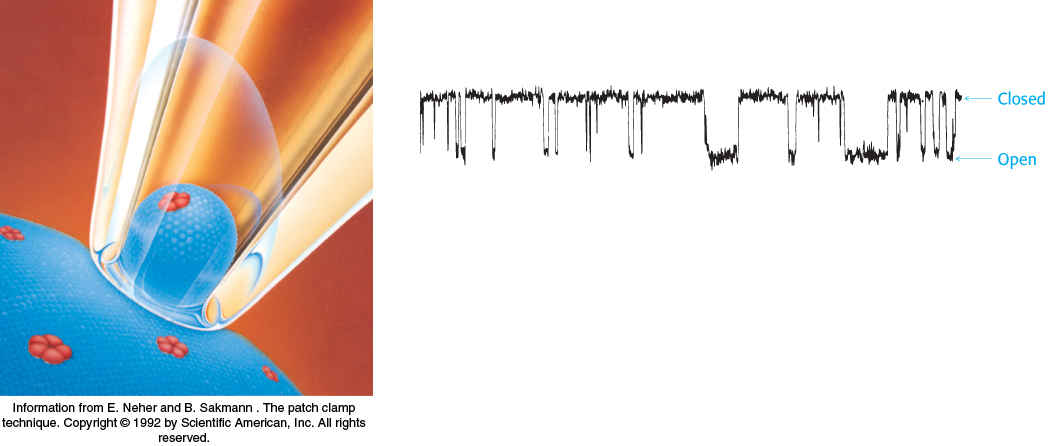
Pumps can establish persistent gradients of particular ions across membranes. Specific ion channels can allow these ions to flow rapidly across membranes down these gradients. These channels are among the most fascinating molecules in biochemistry in their ability to allow some ions to flow freely through a membrane while blocking the flow of even closely related species. The opening, or gating, of these channels can be controlled by the presence of certain ligands or a particular membrane voltage. Gated ion channels are central to the functioning of our nervous systems, acting as elaborately switched wires that allow the rapid flow of current.
Finally, a different class of channel, the cell-
The expression of transporters largely defines the metabolic activities of a given cell type
Each cell type expresses a specific set of transporters in its plasma membrane. This collection of expressed transporters is important because it largely determines the ionic composition inside cells and the compounds that can be taken up from the extracellular environment. In some sense, the specific array of transporters defines the cell’s characteristics because a cell can execute only those biochemical reactions for which it has taken up the necessary substrates.
An example from glucose metabolism illustrates this point. As we will see in Chapter 16, tissues differ in their ability to employ different molecules as energy sources. Which tissues can utilize glucose is largely governed by the expression of members of the GLUT family of homologous glucose transporters. For example, GLUT3 is the primary glucose transporter expressed on the plasma membrane of neurons. This transporter binds glucose relatively tightly so that these cells have first call on glucose when it is present at relatively low concentrations. We will encounter many such examples of the critical role that transporter expression plays in the control and integration of metabolism.