2-6 Ten Principles of Nervous System Function
The balance of the whole nervous system, of the functioning brain, and of individual cells works in concert to produce behavior. Knowing the parts of the nervous system and some general notions about what they do is only the beginning. Learning how the parts work together allows us to proceed to a closer look, in the chapters that follow, at how the brain produces behavior.
Thus far, we have identified 10 principles related to the nervous system’s functioning. Here we elaborate each one. As you progress through the book, review these ideas regularly with an eye toward understanding the concept rather than simply memorizing the principle. Soon you will find yourself applying the principles of function as you encounter new information about the brain and behavior.
Principle 1: The Nervous System Produces Movement in a Perceptual World the Brain Constructs
The nervous system’s fundamental function is to produce behavior, or movement. Movements are not made in a vacuum but are related to objects, places, memories, and myriad other forces and factors. Your mental representation of the world depends on the information sent to your brain. People who are color-
Although we tend to think that the world we perceive is what is actually there, individual realities, both between and within species, clearly are just rough approximations of what is actually present. The brain of each animal species produces a reality that is adaptive for that species to survive. The behavior that the brain produces, in other words, is directly related to the world that the brain has constructed.
Principle 2: Neuroplasticity Is the Hallmark of Nervous System Functioning
Experience alters the brain’s organization, and this neuroplasticity is requisite to learning and memory as well as to survival. In fact, the nervous system stores information only if neural connections change. Forgetting is presumably due to a loss of the connections that represented the memory.
As Experiment 2-1 on page 49 demonstrates, neuroplasticity is a characteristic not just of the mammalian brain; it is found in the nervous system of all animals, even the simplest worms. Nonetheless, larger brains have more capacity for change, and thus their neural organization is likely to show more plasticity.
Detail on plasticity and drug addiction in Section 14-4, on feeling and treating pain in Section 11-4, on epilepsy in Focus 4-1. Section 16-3 details diagnosis and treatment of epilepsy and dementias.
Plasticity can be beneficial in recovering from disorders, such as brain injuries and diseases, as well as in coping with aging. Plasticity also allows the brain to compensate for developmental abnormalities, an extreme example being agenesis of brain structures, as discussed in Research Focus 2-1. Although beneficial in such circumstances, neuroplasticity has drawbacks. Brain analyses of animals given addicting doses of drugs such as cocaine or morphine reveal broad changes in neural connectivity suspected of underlying some maladaptive behaviors related to addiction. Among many other examples of pathological neuroplasticity are those associated with pain, epilepsy, and dementia.
Principle 3: Many Brain Circuits Are Crossed
Most brain inputs and outputs are crossed, that is, serve the opposite side of the body. Each hemisphere receives sensory stimulation from the opposite (contralateral) side of the body and controls muscles on the contralateral side. Crossed organization explains why people who have a stroke or other damage to the left cerebral hemisphere may have difficulty in sensing stimulation to the right side of the body or in moving body parts on the right side. The opposite is true of people whose stroke occurs in the right cerebral hemisphere.
Figure 9-10 illustrates how the human visual system represents the world seen through two eyes as a single perception: both eyes connect with both hemispheres.
A crossed nervous system must somehow join both sides of the perceptual world together. To do so, innumerable neural connections link the brain’s left and right sides. The most prominent connecting cable is the corpus callosum. Its roughly 200 million nerve fibers join the left and right cerebral hemispheres, allowing them to interact.
Four important exceptions to the crossed-
Principle 4: The CNS Functions on Multiple Levels
In simple animals such as worms the spinal cord essentially constitutes the nervous system. More complex animals, such as fishes, have a brainstem as well, and yet more complex animals have also evolved a forebrain. Each new addition to the CNS has added a new level of behavioral complexity without discarding previous levels of control. As animals evolved legs, for example, brain structures evolved to move the legs. Later, the development of independent digit movements required even more brainpower. Thus, new brain areas add new levels of nervous system control. The new levels are not autonomous but rather must be integrated into existing neural systems as refinements and elaborations of the control earlier levels provided.
Multiple levels of function can be seen not only in the addition of forebrain areas to refine brainstem control but also in the forebrain itself. As mammals evolved, they developed an increased capacity to represent the world in the cortex, an ability related to the addition of more maps. The new maps must be related to the older ones, however, and again are simply an elaboration of the perceived sensory world that existed before.
Principle 5: The Brain Is Symmetrical and Asymmetrical
The left and the right hemispheres look like mirror images, but they have some dissimilar features. Cortical asymmetry is essential for integrative tasks, language and body control among them.
Consider speaking. If a language zone existed in both hemispheres, each connected to one side of the mouth, we would actually be able to talk out of both sides of our mouth at once. That would make talking awkward, to say the least. One solution is to locate language control of the mouth on one side of the brain. Organizing the brain in this way allows us to speak with a single voice.
A similar problem arises in controlling body movements in space. We would not want the left and the right hemispheres each trying to take us to a different place. Again, if a single brain area controls this sort of spatial processing, problem solved.
Language control is typically situated on the left side, and spatial functions are typically on the right. The brains of many species have such symmetrical and asymmetrical features. In the bird brain, the control of singing is in one hemisphere, usually on the left side, as is human language. It is likely that birds and humans evolved the same solution independently—
Principle 6: Brain Systems Are Organized Hierarchically and in Parallel
When we consider the multiple levels of CNS function, it becomes apparent that these levels must be extensively interconnected to integrate their processing and produce unified perceptions or movements. The nature of neural connectivity leads to the principle that the brain has both serial (or hierarchical) and parallel circuitry.
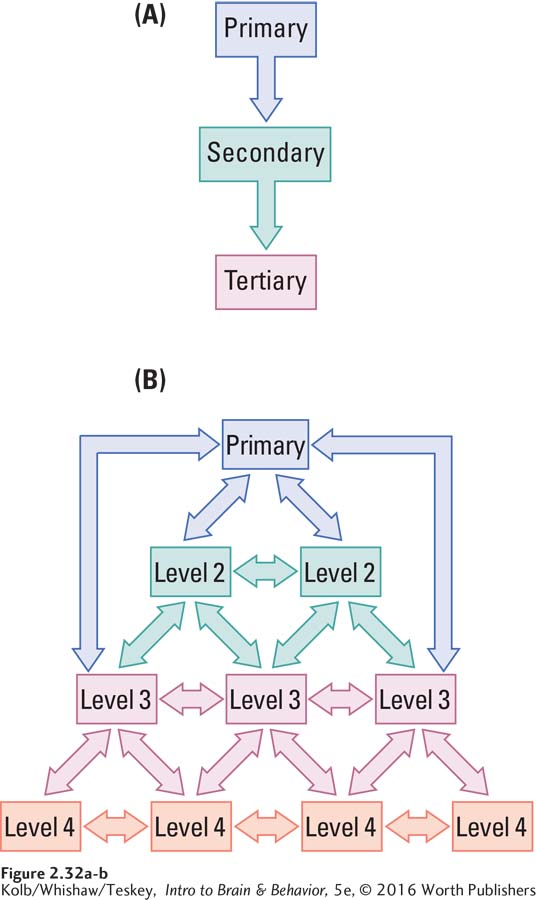
A hierarchical circuit hooks up a linear series of all regions concerned with a particular function. Consider vision. In a serial system, the information from the eyes goes to regions that detect the simplest properties, such as color or brightness. This information is passed along to another region that determines shape, then to another that measures movement, and so on until at the most complex level the information is understood to be, say, your grandmother. Information therefore flows sequentially from simpler to more complex regions in the hierarchy, as illustrated in Figure 2-32A.
However, functionally related brain structures are not always linked linearly. Although the brain has many serial connections, many expected connections are missing. In the visual system, not all cortical regions are connected to one another. The simplest explanation is that the unconnected regions must have widely differing functions.
Parallel circuits operate on a different principle, also illustrated by the visual system. Imagine looking at a car. As we look at a car door, one set of visual pathways processes information about its nature, such as color and shape, whereas another set of pathways processes information about movements such as those necessary to open the door.
These two visual systems are independent of each other, yet they must interact somehow. When you pull the car door open, you do not perceive two different representations—
Figure 2-32B illustrates the information flow in such a distributed hierarchy. If you trace the flow from the primary area to levels 2, 3, and 4, you follow the parallel pathways. And while these multiple parallel pathways are also connected to one another, those connections are more selective than connections in a purely serial circuit.
The brain’s subsystems are organized into multiple parallel pathways, yet our conscious experiences are always unified. As we explore this conundrum throughout the book, keep in mind that your commonsense impressions of how the brain works may not always be correct.
Principle 7: Sensory and Motor Divisions Permeate the Nervous System
The segregation of SNS sensory and motor functions described by the Bell and Magendie law exists throughout the nervous system. Distinctions between motor and sensory functions become subtler in the forebrain.
Sensory and Motor Divisions in the SNS
Review cranial nerve and spinal nerve connections in Figures 2-27 and 2-28.
Spinal nerves are either sensory or motor. Some cranial nerves are exclusively sensory; some are exclusively motor; and some have two parts, one sensory and one motor, much like spinal nerves serving the skin and muscles.
Sensory and Motor Divisions in the CNS
Figures 2-15 through 2-19 illustrate brainstem structures.
The lower brainstem regions—
Distinct sensory nuclei are present in the thalamus, too, although their positions are not segregated, as they are in lower structures. Because all sensory information reaches the forebrain through the thalamus, to find separate nuclei associated with vision, hearing, and touch is not surprising. Separate thalamic nuclei also control movements. Other nuclei have neither sensory nor motor functions but rather connect to cortical areas, such as the frontal lobe, that perform more integrative tasks.
Finally, sensory and motor functions are divided in the cortex in two ways:
Separate sensory and motor cortical regions process a particular set of sensory inputs, such as vision, hearing, or touch. Others control fine movements of discrete body parts, such as the fingers.
The entire cortex is organized around the sensory and motor distinction. As diagrammed in Figure 2-22, layer IV of the cortex always receives sensory inputs, layers V and VI always send motor outputs, and layers I, II, and III integrate sensory and motor operations.
Principle 8: The Brain Divides Sensory Input for Object Recognition and Motor Control
Sensory systems evolved first for controlling motion, not for recognizing things. Simple organisms can detect stimulation such as light and move to or from it. It is not necessary to perceive an object to direct movements toward or away from it. Animals only began to evolve ways of representing their environment as their behaviors became more complex. Animals with a complex brain evolved separate systems for recognizing objects and for moving toward them. The human visual system exemplifies this separation.
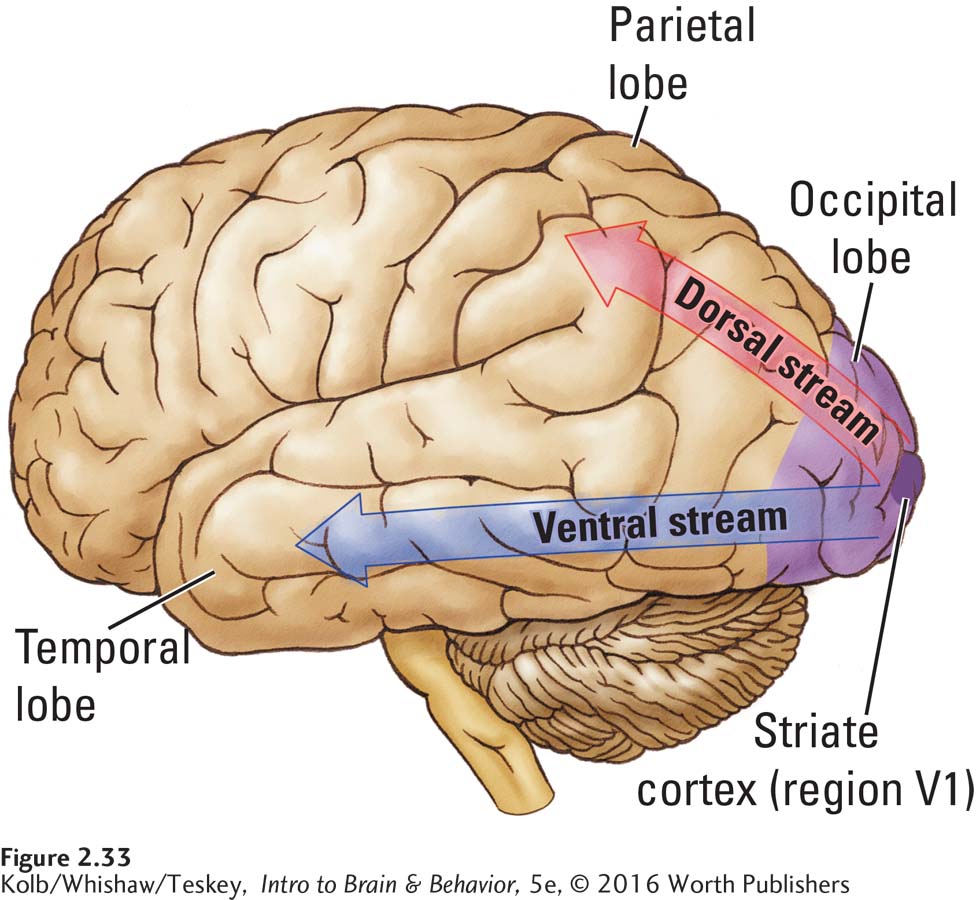
Visual information travels from the eyes to the thalamus to visual regions of the occipital lobe. From the occipital cortex it follows one of two routes: the ventral stream leads to the temporal lobe for object identification, whereas the dorsal stream goes to the parietal lobe to guide movements relative to objects (Figure 2-33). People with ventral stream injuries are blind for object recognition. They cannot distinguish a cup from a spoon. Nevertheless, they shape their hands appropriately when asked to reach for objects that they cannot identify. In contrast, people with dorsal stream injuries can recognize objects, but they make clumsy reaching movements because they do not form appropriate hand postures until they contact objects. Only then do they shape the hand on the basis of tactile information.
Recognizing that perception for movement and perception for object recognition are independent processes has three important implications for understanding brain organization:
The dorsal and ventral visual systems exemplify parallel information processing in the brain.
Although we may think we are aware of our entire sensory world, the sensory analysis required for some movements clearly is not conscious.
Sections 9-2 and 9-3 review evidence that led to understanding the visual streams’ functions and visual information processing.
Unconscious and conscious brain processing underlies an important difference in our cognitive functions. The unconscious movement system is always acting in the present and in response to ongoing sensory input. In contrast, the conscious object recognition system allows us to escape the present and bring to bear information from the past, thus forming the neural basis of enduring memory.
Principle 9: Brain Functions Are Localized and Distributed
A great debate in the history of brain research has concerned what aspects of different functions are actually localized in specific brain regions. Perhaps the fundamental problem is defining a function. Language, for example, includes the comprehension of spoken words, written words, signed words (as in American Sign Language), and even touched words (as in Braille). Language also includes production of words orally, in writing, and by signing, as well as constructing whole linguistic compositions, such as stories, poems, songs, and essays.
Because the function that we call language has many aspects, it is not surprising that these aspects reside in widely separated areas of the brain. We see evidence of this widespread distribution in language-
Figure 14-5 illustrates the extensive distribution of memory areas through the brain.
Memory provides another example of this same distributed pattern. Memories can be richly detailed and can include sensual feelings, words, images, and much more. Like language, then, aspects of memory are located in many brain regions distributed throughout a vast area of the brain.
For Alzheimer neurochemistry, see Section 5-3; for incidence and possible causes, Section 14-3; for treatments, Section 16-4.
Because many functions are both localized and distributed in the brain, damage to a small brain region produces only focal (specific) symptoms. Massive brain damage is required to obliterate some functions. A small injury could impair some aspect of language functioning, such as naming objects, but it would take widespread injury to remove all language abilities. In fact, one characteristic of dementing diseases is that people can endure widespread deterioration of the cortex yet maintain remarkably normal language functions until late stages of the disease. Alzheimer disease is a degenerative brain disorder related to aging that first appears as progressive memory loss and only much later develops into generalized dementia.
Principle 10: The Nervous System Works by Juxtaposing Excitation and Inhibition
We have emphasized the brain’s role in making movements, but we must also recognize that the brain prevents movements. To make a directed movement, such as picking up a glass of water, we must refrain from other movements, such as waving the hand back and forth. In producing movement, then, the brain uses both excitation (increased neural activity) to produce some action and inhibition (decreased neural activity) to prevent other actions.
Brain injury or disease can produce either a loss or a release of behavior by changing the balance between excitation and inhibition. A brain injury in a region that normally initiates speech may render a person unable to talk—
Tourette syndrome and Parkinsonism are dysfunctions of the basal ganglia, which coordinates voluntary movement.
Patients with Parkinson disease may have uncontrollable shaking of the hands because the neural system that inhibits such movements has failed. Paradoxically, they often have difficulty initiating movements and appear frozen because they cannot generate the excitation needed to produce deliberate movements.
The juxtaposition of excitation and inhibition, central to the way the brain produces behavior, can be seen at the level of individual neurons. All neurons evince a spontaneous activity rate that can be either increased (excitation) or decreased (inhibition). Some neurons excite others; some inhibit. Both effects are produced by neuronal communication via specific neurochemicals.
2-6 REVIEW
Ten Principles of Nervous System Function
Before you continue, check your understanding.
Question 1
Many of the brain’s input and output circuits are crossed. In the nervous system, four exceptions to this principle are the __________, the __________, the __________, and the __________.
Question 2
The vertebrate brain has evolved three regions—
Question 3
One aspect of neural activity that resembles the on–
Question 4
Explain this statement: Perception is not reality.
Answers appear in the Self Test section of the book.